| Peripatus Home Page | Updated: 17-Jul-2024 |
AbstractThis page describes the Paleogene Period, including stratigraphy, paleogeography, and famous lagerst‰tten, followed by a sketched outline of some of the major evolutionary events.Keywords: Paleogene, Paleogene biota, fossil record, evolution Introduction |
|
||||||||||
StratigraphyHistorical DevelopmentThe Paleogene System does not have a type section, as such, because it was not originally conceived as a unit in its own right. It was instead “cobbled together” from the earliest three of the seven Cenozoic series (epochs) – Paleocene, Eocene, Oligocene, Miocene, Pliocene, Pleistocene and Holocene – “poetically named for relative abundances of modern forms among the fossil shells” (Ogg et al. 2008, p. 129).
Lower (Cretaceous–Paleogene) BoundarySurely the most widely reported – and the most widely misreported – of all extinction events is that which brought the age of the dinosaurs to an end at the close of the Mesozoic: the “KT” extinction. Almost all of the popular and “lightly technical” literature published within the past couple of decades is filled with comet or meteorite impact theories, all ultimately traceable to the father and son team of Luis and Walter Alvarez. Indeed, there is an overwhelming body of evidence to support the contention that a large extraterrestrial impact did occur at the very end of the Cretaceous; so much so that the Cretaceous-Paleogene boundary is now defined by two impact proxies. The GSSP for the base of the Danian Stage (= Cretaceous-Paleogene boundary) was established in 1996 in the clay layer containing the famous “K/T” iridium anomaly and nickel-rich spinels at a section of Oued Djerfane, 8 km west of El Kef, in Tunisia (see Molina et al. 2006).
Upper (Paleogene–Neogene) BoundaryThe Paleogene-Neogene boundary is coincident with the base of the Aquitanian Age which in turn defines the base of the Miocene Epoch. The Aquitanian GSSP is formally defined in the Lemme-Carrosio section in northern Italy, located at the 35 m level as measured downward from the top of the section. The horizon corresponds closely with the first appearance of the calcareous nannofossil Sphenolithus capricornutus and with the C6Cn.2n(o) reversal boundary (after Gradstein et al. 2012, p. 925-926).
ChronologyThe Paleogene Period extends from 66.0 Ma to 23.03 Ma (Cohen et al. 2015).
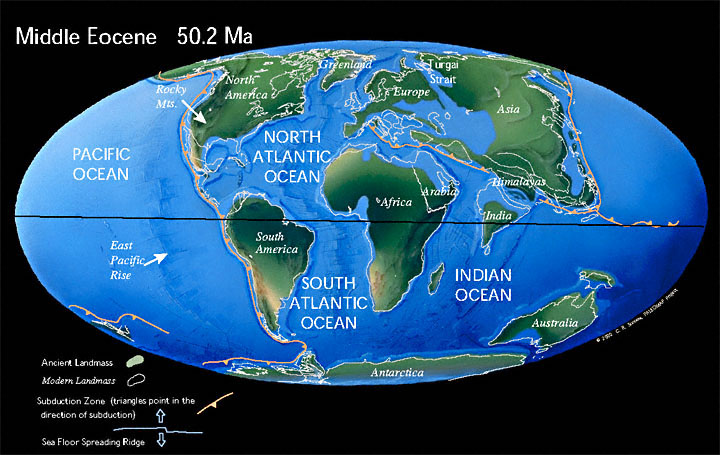
What happened to the Tertiary?A great deal of the older, particularly European, literature subdivides the Cenozoic into the Teriary and Quaternary Periods, the Tertiary Period comprising the Paleocene through Pliocene Epochs. In fact, this scheme has a very long history, dating back to the earliest attempts to subdivide geological time in the mid-1700s. An alternative subdivision into Paleogene and Neogene was introduced by Hörnes in 1853, and took root, most strongly in America. Initially, the Neogene extended to the present day, i.e., it incorporated the Quaternary Period also. After some indecision, the ICS eventually adopted a three-fold subdivision of the Cenozoic into Paleogene, Neogene and Quaternary periods. The Paleogene Period comprises the Paleocene, Eocene, and Oligocene epochs, the Neogene comprises the Miocene and Pliocene epochs, and the Quaternary encompasses the Pleistocene, Holocene, and assuming to is eventually ratified, Anthropocene. PaleogeographyMajor Tectonic Events“In another outcome of middle Eocene tectonic events the sedimentary basins of southern Australia formed – or some were rejuvenated – in a shakeup of the northern margin of the AAG [the Australo-Antarctic Gulf, an initially blind seaway formed as an eastward-propagating fracture between the future Antarctica and Australia when Gondwana began to break up]. There was some uplift, enough to stimulate the developing of a continent-wide system of incised channels draining into the AAG and also northward into the Lake Eyre Basin, for the modern endorheic (closed) drainage of central Australia dates from that time. Ö Embedded in the geohistorical and biohistorical culture of southern Australia is the belief that the climate changed due to the continent migrating towards the equator. This is part of the whole truth, but it must be added that global oceanic isotherms at times oscillated through tens of degrees of latitude whilst seafloor spreading was shifting the continent ~100 km (Figure 1). The early Paleogene tropics were reaching towards southern Australia long before Australia approached the Neogene tropics” (McGowran & Hill 2015, p. 23). |
|||||||||||
Land and SeaIn the Southern Hemisphere, one of the major paleogeographic events was the opening and deepening of the Drake Passage and Tasmanian Gateway (in whichever order) allowing the Antarctic Circumpolar Current to form. Initially it was generally believed that this resulted in the “thermal isolation” of Antarctica, leading to the development of the continental scale glaciation we see today. More recently this view has been challenged, and it is now quite controversial.The essentials of the Tasmanian Gateway were sketched out by Kennett et al. in 1974 but the following brief description is taken from a later paper, Kennett & Exon 2004: “(1) During the Eocene, before the gateway opening, the absence of circum-Antarctic circulation allowed flow of warmer surface currents to high southern latitudes, thus warming Antarctica; (2) Gateway opening at the Eocene/Oligocene boundary allowed initial development of the Antarctic Circumpolar Current which immediately disrupted southward flow of warmer waters in the Indo-Pacific sector of the Southern Ocean, in turn triggering major cooling and Antarctic glacial development. A major climate shift at this time is indicated by a marked positive oxygen isotopic shift in the earliest Oligocene [Shackleton and Kennett, 1975; Miller et al. 1987; Zachos et al. 2001]. The magnitude and relative suddenness of the climate change is striking [Berggren and Prothero, 1992]. Strong positive feedbacks were almost certainly involved, including albedo changes due to expansion of snow and ice Ö, atmospheric green house gases Ö and Southern Ocean biological productivity Ö” (p. 347). |
(1) |
||||||||||
ClimateThe most significant climatic event in the Paleogene, and one of the most significant events of the whole Phanerozoic, was the Paleocene–Eocene Thermal Maximum (PETM). This was a geologically brief (probably lasting fewer than 200,000 years) global warming event. Brief temperature “spikes” like this are called hyperthermals. As the name suggests, it occurred at or very close to the boundary between the Paleocene and Eocene epochs, approximately 56 million years ago, at a time when the climate was already warm (Kennett and Stott 1991; Zachos et al. 2001, 2008; Zeebe & Lourens 2019). During the PETM, global temperatures rapidly increased by 5–8°C within 20 ka (Wing et al. 2005; Weijers et al. 2007; McInerney & Wing 2011).“Early Paleogene southern Australia, which comprises the north shore of the Australo-Antarctic Gulf, was covered in the equivalent of extant wet-tropical lowland vegetation at 60–65∞S, most richly during the Early Eocene Climatic Optimum. Cooling at ~50 Ma triggered the spread of Nothofagus warm-temperate rainforests. Cooling reversed at the Khirthar transgression at ~42 Ma with extensive neritic carbonate seas, diverse rainforest biomes and massive coals on the continental margins. Ö Terrestrial floras were still wet-temperate at the continental margins, but during the Oligocene various plants, already coping with low nutrients from extensive deep weathering in the Paleogene, had to cope with cooling, sudden drops in global CO2 levels and a drying-out in the continental interior. The rapid expansion of the Antarctic ice sheet in the early Oligocene had biotic impacts and responses, but not the dramatic impact of the subsequent expansion at ~14 Ma” (McGowran & Hill 2015, Abstract). Paleontology
Major TaxaAnurans (frogs and toads)Recent phylogenetic analysis suggests that all extant families and subfamilies containing arboreal species originated after the Cretaceous, suggesting that new ecological opportunities shaped anuran diversification as forests rebounded after ... vegetation loss at the K–Pg extinction event” (Xing et al. 2018, p. 1). Mammalian RadiationBoth abundance and diversity of mammals were reduced severely by the Cretaceous/Paleogene mass extinction event (e.g. in the Hell Creek assemblage, only one out of 28 mammal species survived; Ward 2000, p. 173) with fewer taxa known from the Paleocene than from the Cretaceous. During the Cretaceous, mammals were mainly small, terrestrial-to-arboreal insectivores with low ecological disparity (Goswami 2012, Grossnickle & Polly 2013, Halliday et al. 2017), albeit with a few notable exceptions (Luo 2007). The analysis by Meredith et al. (2011) suggests that “only 29 to 32 mammalian lineages, nearly all of which are stem branches leading to extant orders, may have crossed the KPg boundaryÖ. This inference is consistent with the long-fuse model of mammalian diversification ... which postulates interordinal diversification in the Cretaceous, followed by intraordinal diversification that is mostly restricted to the Cenozoic ..., although conflicts do remain with the Cretaceous eutherian fossil record” (p. 523). Whereas early placentals probably lived in the Mesozoic (although no Cretaceous eutherian mammal has been unambiguously resolved within the placental crown; Wible et al. 2009, Goswami et al. 2011, Halliday et al. 2017), the crown group radiation is thought to have occurred after the end of the Cretaceous (e.g., O’Leary et al. 2013, Halliday et al. 2017). Paleogene placental mammals include the first large-bodied herbivores, specialised carnivores, and later, radiations of gliding, flying, and fully aquatic organisms, with a corresponding increase in diversity (Darroch et al. 2014, Halliday et al. 2017). “By far the largest component of the mammalian biota in the Paleocene is the collection of ‘archaic ungulates’ known as ‘condylarths’. While this grouping is almost certainly an anachronistic grade of largely terrestrial, bunodont, herbivorous-to-omnivorous mammals, there are several well-defined families which fall within ‘Condylarthra’” (Halliday et al. 2017, p. 525). The earliest stem carnivorans, the group including cats, dogs, bears and others, are the genera Ravenictis and Pristinictis, known from the earliest Paleocene, and afrotherians (the group comprising elephants, dugongs, aardvarks, among others) from the Middle Paleocene. The first chiropteran (bat) fossils – already quite highly derived, capable of true flight although not echolocation – are known from the famous, Early Eocene Green River Formation of Wyoming. The earliest lagomorphs (rabbits, hares, etc.) are known from the mid Eocene of China. (After Halliday et al. 2017.) The diversity and range of mammals appears to have greatly increased after the Paleocene/Eocene boundary (about 55 million years ago), and new groups appeared on continents throughout the Northern Hemisphere. On the basis of primarily phylogenetic analyses, Asia has been suggested as a likely centre of origin (Bowen et al. 2002, p. 2028). A study by Bininda-Emonds et al. (2007) examined relations and divergence times among all living mammalian families and concluded that there was a dramatic upturn in mammalian diversification rates during the Eocene, ~55 to 50 million years ago. This idea has been challenged, however (see Meredith et al. 2011). Early Primate EvolutionThe closest living relatives of primates may be the Dermoptera (colugos, arboreal gliding mammals native to Southeast Asia; suggested by genetic studies) or the clade of Dermoptera plus Chiroptera (bats; suggested by morphology). Any common ancestor to the three groups must have lived in the Cretaceous. “If the Cretaceous Indian genus Deccanolestes is, as some have suggested, closely related to purported euarchontans, such as nyctitheres ... or adapisoriculids ... then Deccanolestes would represent a Cretaceous occurrence of a euarchontan” (the group containing rodents and primates, among others; Halliday et al. 2017, p. 531, and references therein). The various groups of “stem” primates (for our purposes, ancestral primates which have not yet evolved all the characteristics of modern primate taxa) are collectively known as plesiadapiforms. This is not a “proper” taxonomic group; the relationships between its various members are poorly known and the group as a whole is probably paraphyletic. Of these composite taxa, the geologically oldest and most primitive known family is the Purgatoriidae, which includes the genus Purgatorius. “A Cretaceous origin A Cretaceous origin of Pan-Primates has long been hypothesized by palaeontologists, in part based on the initial description of Purgatorius, including P. ceratops, which until more recently ... was considered latest Cretaceous in age.... A consensus later emerged that despite the lack of unambiguous records of Cretaceous plesiadapiforms, a pre-KPB origin of Pan-Primates was still within the realm of possibility.... Fox & Scott [Fox & Scott 2011] recently speculated that the early (Pu2) occurrence and derived characteristics of Purgatorius coracis imply that the ancestral purgatoriid was from the Late Cretaceous. Fossils reported here (i) derive from an older (Pu1) locality, less than 208 kyr and most likely to within 105–139 kyr post-KPB, and (ii) represent two sympatric species of Purgatorius, each with a uniquely accumulated suite of dental specializations that evolved following divergence from a common ancestor. These data provide even stronger evidence that the origin of plesiadapiforms, and in turn Pan-Primates, Euarchonta and Placentalia extends back into the Late Cretaceous” (Mantilla et al. 2021, p. 7). The primate record, generally, and the human record in particular, is very incomplete. Probable primate ancestors – genera such as Purgatorius, Plesiadapis and Phenacolemur – date from the earliest Paleogene, approximately 66-65 Ma. The oldest known true primates occur about 55 Ma (near the Paleocene/Eocene boundary). Lagerst‰ttenDominican Amber: ?Eocene to Miocene (15-45 Ma); Dominican Republic; small animals, mostly arthropds, and plant fragments preserved in amber; Poinar & Poinar 1999Grube Messel Shale: Eocene Frankfurt, Germany; lacustrine (lake deposits); fossil plants, vertebrates and insects; Schaal & Ziegler 1992 Monte Bolca (Mt. Bolca): 52 Ma Eocene Near Verona, Italy; tropical marine lagoon. Exceptional preservation of fishes (>200 different kinds), plants, leaves and rare insects. Known since the 17th century. Also see the Musei della Lessinia web page (in Italian). [With thanks to Giorgio Bertoni for this information.] Green River Formation: Eocene Wyoming; lacustrine (lake deposits); fossil fish (~18 different kinds) and other vertebrates; Grande 1984 Insect Limestone: Eocene, Isle of Wight; includes numerous plant and animal taxa but especially well-known for arthropods, Ross & Self 2014
New Zealand OccurrencesAs Zealandia continued to rift away from Gondwana, the new microcontinent underwent post-rift lithospheric cooling and slowly subsided. The subsidence resulted in extensive marine transgression, with consequent reduction in land area, frequently expressed in the geological record by terrestrial sequences such as coal measures overlain by marine sediments. By the Middle Eocene, “the reduced Zealandia landmass had extensive coastal plains and predominantly fine-grained clastic sediments and carbonates were accumulating in marine sedimentary basins offshore” (Edbrooke 2017, p. 31). Subsidence of the Zealandia microcontinent reached a maximum between about 35 and 25 Ma, in the Oligocene. This interval of “maximum drowning” saw widespread deposition of limestones as the clastic sediment supply diminished due to the dwindling of emergent land. Some authors have gone so far as to suggest that Zealandia became completely submerged at some time in the late Oligocene, and there is lively on-going debate about this hypothesis. The principal counter-argument is that the diverse and highly endemic present-day New Zealand biota is unlikely to have been able to develop de novo since the end of the Oligocene. ReferencesBerggren, W.A.; Prothero, D.R. 1992: Eocene-Oligocene climatic and biotic evolution: an overview. In Prothero, D.R.; Berggren, W.A. (ed.) 1992: Eocene-Oligocene climatic and biotic evolution. Princeton University Press: 1-28. Bininda-Emonds, O.R.P.; Cardillo, M.; Jones, K.E.; MacPhee, R.D.E.; Beck, R.M.D.; Grenyer, R.; Price, S.A.; Vos, R.A.; Gittleman, J.L.; Purvis, A. 2007: The delayed rise of present-day mammals. Nature 446: 507-512. Bowen, G.J.; Clyde, W.C.; Koch, P.L.; Ting, S.; Alroy, J.; Tsubamoto, T.; Wang, Y.; Wang, Y. 2002: Mammalian Dispersal at the Paleocene/Eocene Boundary. Science 295: 2028-2029. Cohen, K.M.; Finney, S.C.; Gibbard, P.L.; Fan, J.X. 2015: The ICS international chronostratigraphic chart v 2015/01. Episodes 36: 199-204. Darroch, S. A. F.; Webb, A. E.; Longrich, N.; Belmaker, J. 2014: Palaeocene-Eocene evolution of beta diversity among ungulate mammals in North America. Global Ecology and Biogeography 23: 757-768. Edbrooke, S.W. 2017: The geological map of New Zealand. GNS Science Geological Map 2: 1-183. Fox R.C.; Scott C.S. 2011: A new, early Puercan (earliest Paleocene) species of Purgatorius (Plesiadapiformes, Primates) from Saskatchewan. Canadaian Journal of Paleontology 85: 537-548. Goswami, A. 2012: A dating success story: genomes and fossils converge on placental mammal origins. EvoDevo 3: 4. Goswami, A.; Prasad, G.V.R.; Upchurch, P.; Boyer, D.M.; Seiffert, E.R.; Verma, O.; Gheerbrant, E.; Flynn, J.J. 2011: A radiation of arboreal basal eutherian mammals beginning in the late Cretaceous of India. Proceedings of the National Academy of Sciences of the United States of America 108: 16333-16338. Gradstein, F.M.; Ogg, J.G.; Schmitz, M.D.; Ogg, G.M. 2012: The Geologic Time Scale 2012. Elsevier 1-2. Grande L. 1984: Paleontology of the Green River Formation. Bulletin of the Geological Survey of Wyoming, no. 63: 1-333. Grossnickle, D.M.; Polly, P.D. 2013: Mammal disparity decreases during the Cretaceous angiosperm radiation. Proceedings of the Royal Society B: Biological Sciences 280. Halliday, T.J.D.; Upchurch, P.; Goswami, A. 2017: Resolving the relationships of Paleocene placental mammals. Bioogical Reviews 92: 521-550. Hörnes, M. 1853: Mitteilung an Prof. Bronn gerichtet: Wien, 3. Okt., 1853. Jahrbuch f¸r Mineralogie, Geognosie, Geologie und Petrefakten-Kunde: 806-810. Kennett, J.P.; Exon, N.F. 2004: Paleoceanographic evolution of the Tasmanian Seaway and its climatic implications. In Exon, N.F.; Kennett, J.P.; Malone, M.J. (ed.) 2004: The Cenozoic Southern Ocean: Tectonics, Sedimentation, and Climate Change Between Australia and Antarctica. AGU Geophysical Monograph Series 151: 345-367. Kennett, J.P.; Houtz, R.E.; Andrews, P.B.; Edwards, A.R.; Gostin, V.A.; Hajos, M.; Hampton, M.A.; Jenkins, D.G.; Margolis, S.V.; Ovenshine, A.T.; Perch-Nielsen, K. 1974: Development of the Circum-Antarctic Current. Science 186: 144-147. Kennett, J.P.; Stott, L.D. 1991: Abrupt deep-sea warming, palaeoceanographic changes and benthic extinctions at the end of the Palaeocene. Nature 353: 225-229. Luo, Z.X. 2007: Transformation and diversification in early mammal evolution. Nature 450: 1011-1019. Mantilla, G.P.W.; Chester, S.G.B.; Clemens, W.A.; Moore, J.R.; Sprain, C.J.; Hovatter, B.T.; Mitchell, W.S.; Mans, W.W.; Mundil, R.; Renne, P.R. 2021: Earliest Palaeocene purgatoriids and the initial radiation of stem primates. Royal Society Open Science 8 (210050): 1-10. McGowran, B.; Hill, R.S. 2015: Cenozoic climatic shifts in southern Australia. Transactions of the Royal Society of South Australia 139 (1): 19-37. McInerney, F.A.; Wing, S.L. 2011: The Paleocene-Eocene thermal maximum: a perturbation of carbon cycle, climate, and biosphere with implications for the future. Annu. Rev. Earth Planet. Sci. 39: 489-516. Meredith, R.W.; Janečka, J.E.; Gatesy, J.; Ryder, O.A.; Fisher, C.A.; Teeling, E.C.; Goodbla, A.; Eizirik, E.; Simão, T.L.L.; Stadler, T.; Rabosky, D.L.; Honeycutt, R.L.; Flynn, J.J.; Ingram, C.M.; Steiner, C.; Williams, T.L.; Robinson, T.J.; Burk-Herrick, A.; Westerman, M.; Ayoub, N.A.; Springer, M.S.; Murphy, W.J. 2011: Impacts of the Cretaceous terrestrial revolution and KPg extinction on mammal diversification. Science 334: 521-524. Miller, K.G.; Fairbanks, R.G.; Mountain, G.S. 1987: Tertiary oxygen isotope synthesis, sea-level history, and continental margin erosion. Paleoceanography 2: 1-19. Molina, E.; Alegret, L.; Arenillas, I.; Arz, J.A.; Gallala, N.; Hardenbol, J.; von Salis, K.; Steurbaut, E.; Vandenberghe, N.; Zaghbib-Turki, D. 2006: The Global Boundary Stratotype Section and Point for the base of the Danian Stage (Paleocene, Paleogene, ìTertiaryî, Cenozoic) at El Kef, Tunesia - Original definition and revision. Episodes 29: 263-273. Ogg, J.G.; Ogg, G.; Gradstein, F.M. 2008: The concise geologic time scale. Cambridge University Press: 1-177. O’Leary, M.A.; Bloch, J.I.; Flynn, J.J.; Gaudin, T.J.; Giallombardo, A.; Giannini, N.P.; Goldberg, S.L.; Kraatz, B.P.; Luo, Z.; Meng, J.; Ni, X.; Novacek, M.J.; Perini, F.A.; Randall, Z.S.; Rougier, G.W.; Sargis, E.J.; Silcox, M.T.; Simmons, N.B.; Spaulding, M.; Velazco, P.M.; Weksler, M.; Wible, J.R.; Cirranello, A.L. 2013: The placental mammal ancestor and the post-K-Pg radiation of placentals. Science 339: 662-667. Poinar, G. Jr.; Poinar, R. 1999: The amber forest: A reconstruction of a vanished world. Princeton: 1-239. Ross, A.J.; Self, A. 2014: The fauna and flora of the Insect Limestone (late Eocene), Isle of Wight, UK: introduction, history and geology. Transactions of the Royal Society of Edinburgh 104: 233-244. Schaal, S.; Ziegler, W. (ed.) 1992: Messel. An insight into the history of life and of the Earth [English translation]. Clarendon Press: 1-322. Shackleton, N.J.; Kennett, J.P. 1975: Paleotemperature history of the Cenozoic and the initiation of Antarctic glaciation: oxygen and carbon isotope analyses in DSDP sites 277, 279, and 281. In Kennett, J.P., Houtz, R.E., et al. 1975: Initial reports of the Deep Sea Drilling Project. US Government Printing Office, Washington 29: 743-755. Ward, Peter D. 2000: Rivers in Time. Columbia University Press: 1-320. Weijers, J.W.; Schouten, S.; Sluijs, A.; Brinkhuis, H.; Sinninghe Damste, J.S. 2007: Warm arctic continents during the Palaeocene-Eocene thermal maximum. Earth Planet. Sci. Lett. 261: 230-238. Wible, J. R.; Rougier, G. W.; Novacek, M. J.; Asher, R. J. 2009: The eutherian mammal Maelestes gobiensis from the late cretaceous of mongolia and the phylogeny of Cretaceous Eutheria. Bulletin of the American Museum of Natural History 327: 1-123. Wing, S.L.; Harrington, G.J.; Smith, F.A.; Bloch, J.L.; Boyer, D.M.; Freeman, K.H. 2005: Transient floral change and rapid global warming at the Paleocene-Eocene boundary. Science 310: 993-996. Xing, L.; Stanley, E.L.; Bai, M.; Blackburn, D.C. 2018: The earliest direct evidence of frogs in wet tropical forests from Cretaceous Burmese amber. Nature Scientific Reports 8 (8770): 1-8. Zachos et al. 2001: Science 292: 274. Science 292: 274. Zachos, J.; Dickens, G.; Zeebe, R. 2008: An early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics. Nature 451: 279-283. Zeebe, R.E.; Lourens, L.J. 2019: Solar system chaos and the Paleocene-Eocene boundary age constrained by geology and astronomy. Science 365: 926-929. |
| Peripatus Home Page |
Hits counted from 22 Feb 2018:
My Traffic Estimate