| Peripatus Home Page | Updated: 31-Jan-2024 |
AbstractThe Cambrian was a time of mild, equitable climate, when high sea levels covered extensive continental margins. Sediments deposited on the margins of one of the Cambrian continental masses, Avalonia, now form the type section in Wales. Modern dating techniques have shown that the Cambrian was of shorter duration than previously thought: the currently accepted duration is ~56 Ma, from 541 to ~485 Ma ago. The period occupies a special place in the study of paleontology, because it is in the sediments of Cambrian age that fossils “suddenly” become common for the first time. This effect has come to be known as the “Cambrian explosion.” Most modern phyla capable of leaving fossils are known from the Cambrian, though, numerically, fossil collections from this interval are dominated by arthropods. Best known of these are certainly the trilobites. Much of our knowledge of early metazoan evolution has come from the study of three remarkable lagerstätten: Sirius Passet in northern Greenland, Chengjiang in southern China, and the Burgess Shale of western Canada. Extinction rates were generally high throughout the Cambrian, with two major peaks qualifying as mass extinctions.Keywords: stratigraphy, Cambrian Period, Cambrian biota, fossil record, evolution, metazoan radiation, extinction, Burgess Shale, Chengjiang IntroductionThe Cambrian Period is the earliest period in the Paleozoic Era, the latter name meaning “ancient life” because this interval is characterised by the presence of distinctive fossils – such as the famous trilobites – which look very unlike more modern creatures. Near the beginning of the Cambrian interval, life began to rapidly evolve from earlier cryptic forms, which were very poorly known until late in the twentieth century, into larger and diverse forms that have been known as fossils since the early 1800s. Famously, Charles Darwin was discomforted by the fact that the earliest fossils known in his time were already complex, obviously highly evolved, requiring earlier, simpler forms to be hypothesised. Precambrian fossils are no longer hypothetical: hundreds or thousands of different kinds are now known; many well-known (read more). However, it does really seem as if something happened in the late Early to early Middle Cambrian. Prior to then, the fossil record is sparse and the fossils themselves mostly very strange and unfamilar, whereas, in the Chengjiang deposit, for example, a vast number and diversity of different organisms is apparent, including a great many of the major groups of animals living today. This phenomenon is known colloquially as the “Cambrian explosion”. The Cambrian Period was erected by Adam Sedgwick and Roderick Murchison (1835) who named it after the Roman name for Wales, Cambria, where they were mapping. Although the Welsh rocks were first to bear the name Cambrian, the oldest beds of the system are not well-developed there. The best sections, or at least the best studied, are to be found on the Siberian Platform and in China, and for many years the Siberian rock units, particularly, have been widely adopted as de facto subdivisions of the system. However, good sections are increasingly becoming known from Australia, Canada, and elsewhere. Two particularly famous fossil localities, Chengjiang (south of Kunming, China) and Yoho National Park (in the British Columbian Rocky Mountains), are known to almost everyone with an interest in paleontology. These have yielded thousands of spectacularly well-preserved fossils, revealing more about middle Cambrian marine life than we know about many younger times. |
|
||||||||||||||||||||||||
StratigraphyHistorical Development“Adam Sedgwick named the Cambrian in 1835. He derived the name from ‘Cambria,’ the Roman name for Wales, site of the type area, in which shales and sandstones make up a section about two miles [~3 km] thick. Though these rocks are strongly folded and faulted, some are fossiliferous” (Stirton 1967, p. 68). In the 1830s, Sedgwick and Sir Roderick Impey Murchison, commenced work on the stratigraphy of northern Wales. Sedgwick began at the bottom of the section and Murchison at the top. Sedgwick named his sequence of rocks the Cambrian and Murchison applied the name Silurian to the generally more fossiliferous upper formations (Sedgwick & Murchison 1835). Eventually their sections overlapped, each claiming some of the same rocks for their systems, in what became a widespread and lengthy controversy until, forty four years later, Charles Lapworth introduced the name Ordovician for the disputed sequence. The type locality is in western Wales and is defined mainly by lithology.
Lower (Ediacaran–Cambrian) Boundary |
|||||||||||||||||||||||||
|
Since 1947, when H.E. Wheeler initiated debate with the suggestion that the Precambrian-Cambrian boundary should be based upon the first appearance of trilobites, much has ensued. Progress has largely been facilitated by the International Geological Congress (IGC) and the establishment in 1960 of a Subcommission on Cambrian Stratigraphy. The classical idea of placing the boundary at an unconformity has been displaced by the search for monofacial, continuous deposition sequences across the boundary, with the view to selecting a stratotype. The search itself produced a wealth of data from around the world – including the Palaeotethyan Belt, Siberian Platform, and England – eventually focusing upon south-east Newfoundland. Prior to 1990, the boundary was generally placed at the base of the Tommotian Stage, a rock unit known from the Siberian Platform. However, in 1991 the International Subcommission on Cambrian Stratigraphy made a decision to draw the base on the Cambrian at the first appearance datum (FAD) of the distinctive horizontal burrow ichnofossil, Treptichnus (formerly Phycodes and Trichophycus) pedum (Seilacher 1955; Fig. 1), in the reference section at Fortune Head. This horizon correlates with the base of the Siberian Nemakit-Daldynian Stage, ~13 Ma earlier than the Tommotian (included within the Vendian System in some older literature). Both of the Siberian stages have since been abandoned by the ISCS, however. Presently, the early Cambrian is represented by the Terreneuvian Series, which is divided into the Fortunian Stage (basal) and “Stage 2”. This is unlikely to be the final word on the matter. |
(1) Fig. 1: The horizontal burrow trace fossil, Treptichnus (formerly Phycodes and Trichophycus) pedum defines the lower boundary of the Cambrian in the reference section at Fortune Head, southeastern Newfoundland. [Image courtesy of Dr. Gerd Geyer, Institut für Paläontologie, Bayerische Julius-Maximilians-Universität, Würzburg, Germany.] |
||||||||||||||||||||||||
|
Upper (Cambrian–Ordovician) BoundaryThe Cambrian was characterised by generally high extinction rates; at least two events are usually ranked among the great Phanerozoic “mass extinctions”. The later of these two events occured at or near the end of the period and notably decimated the trilobites. Steadily accumulating data has led to changing opinions as to where the Cambrian-Ordovician boundary should be formalised. “Traditional concepts of the Cambrian-Ordovician boundary [were] based on the occurrence of the graptolite Dictyonema (formerly Rhabdinopora) flabelliforme (Eichwald 1840), which has a limited regional distribution. Strata with Dictyonema flabelliforme are often difficult to correlate precisely so that different species (and subspecies) of conodonts were favoured for definition of the Cambrian-Ordovician boundary. The Cambrian-Ordovician Boundary Working Group finally decided in 1998 by majority that the base of the Ordovician should be placed at the base of the zone with Iapetognathus fluctivagus, which approximates the Cordylodus lindstromi Zone, the Dictyonema flabelliforme Zone and the FAD of the trilobite Jujuyaspis” (this text formerly available on the International Subcommission on Cambrian Stratigraphy web page). The GSSP for this boundary was chosen at the Green Point section, Newfoundland, approved by the International Commission on Stratigraphy in December 1999 and ratified by the IUGS in January 2000. By a unanimous vote taken in February 2000, the Subcommission selected the name Tremadocian for the lowest stage of the Ordovician System.
Chronology |
|||||||||||||||||||||||||
|
Subdivisions of the Cambrian System were for a long time based on the rock units of the Siberian Platform, which became well known everywhere and are still often used today (Fig. 2).
The International Commission on Stratigraphy (ICS) appears to be largely abandoning the Siberian units; many epoch- and age-level subdivisions of the Cambrian are yet to be defined but those that are based on rock sequences in Canada and China. Until 1991, when the base of the Cambrian was finally formally defined, the base of the system was most often taken as the base of the Tommotian Stage (and one commonly still finds that in older literature). The new definition added the Nemakit-Daldynian (or Manykaian) below the Tommotian, enlarging the system by approximately a half. Notwithstanding the extension of the Cambrian definition in 1991, geochronological studies carried out during the past few decades have provided new absolute age estimates which actually reduced the understood duration of the Cambrian. In general, the time interval attributed to the unit has shrunk because the reduction in the understood age of the Cambrian-Ordovician boundary has been exceeded by a larger reduction in the understood age of the Precambrian-Cambrian boundary. Some influential published estimates at approximately decadal intervals are:
The International Subcommission on Cambrian Stratigraphy has settled on an age of 541 Ma for the Precambrian-Cambrian boundary, and 485.4 ± 1.9 Ma for the Cambrian-Ordovician boundary (Cohen et al. 2015, 2017). These are the values adopted throughout this site. |
(2) Fig. 2: International and Siberian Platform timescales for the Cambrian (after Ogg et al. 2016). |
||||||||||||||||||||||||
PaleogeographyMajor Tectonic EventsThe Precambrian supercontinent variously known as Proto-Pangea, Ur-Pangea, and Rodinia, formed ~1,000 Ma from the amalgamation of three or four pre-existing continents, in an event known as the Grenville Orogeny. Perhaps beginning ~700 Ma, but protracted over many millions of years, Rodinia began breaking up into three major blocks: West Gondwana, East Gondwana, and Laurasia. Subsequently – perhaps ~540 Ma, near the beginning of the Cambrian – West and East Gondwana merged in the mountain-building event known as the Pan-African Orogeny. (After Rogers 1996.) |
|||||||||||||||||||||||||
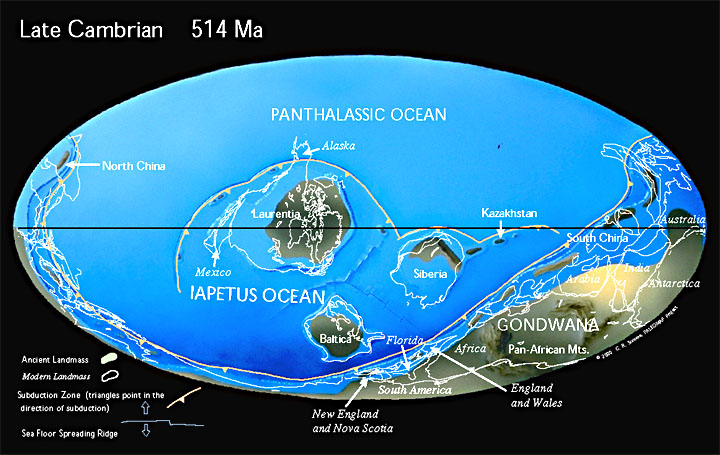
Land and SeaGondwana, comprising much of what is now India, Australia, New Zealand, Antarctica, Africa and South America, was a super-continent stretching from ~35şN almost to the South Pole.North America and parts of Scotland were united in the subequatorial continent of Laurentia, surrounded by calm seas allowing the accumulation of extensive limestone deposits in marginal basins (Fig. 3). Approximately 505 Ma ago, a mudslide on the western continental margin of Laurentia, in what is now eastern British Columbia, buried the famous Burgess Shale fauna. Other land masses included Baltica, comprising much of modern western Europe, also near the South Pole, and Siberia, which occupied equatorial latitudes. The Iapetus Ocean stretched between Laurentia and Baltica; present day Greenland and Scandinavia occupying the continental margins of Laurentia and Baltica, respectively. The north and south of present day China were separated, lying beneath shallow seas in northern tropical latitudes. The evidence concerning north Africa in Early Cambrian times is equivocal: paleomagnetic studies indicate a position near the South Pole, adjacent to the south of Gondwana, yet the faunas, which include coral reefs, are tropical. Avalonia, comprising Wales and parts of England, Newfoundland, New England and Nova Scotia, was also submarine at this time, and near the South Pole. |
(3) |
||||||||||||||||||||||||
ClimateAlthough ice ages both preceded and followed the Cambrian, there is no direct evidence for significant glaciation during the period, and the conventional view is that, in general, the global climate was probably warmer and more uniform than it is today.Separate landmasses arising from the breakup of Rodinia were distributed from low (tropical) and southern temperate latitudes (North America and Siberia) or somewhat to the south (Baltica). Most of Rodinia remained in the supercontinet Gondwana, however, which stretched from the tropics to the south pole, where it is likely conditions were cooler than elsewhere. A 2018 study of oxygen isotopes from phosphatic fossils extracted from the Comley Limestones, Shropshire, on the Avalonia microcontinent (Hearing et al. 2018), concluded the limestones were deposited in a shallow marine setting reflecting high southern latitude sea surface temperatures of 20 to 25C. However, as Babcock et al. (2015, p. 12) point out, “interrelated, cyclic, physical, chemical, and biotic effects during the Cambrian Period produced distinctive, time-specific global patterns of facies. These changes include abrupt episodes of sea level rise and incursion of cooler water onto the shelves of Cambrian continents. … The Cambrian Period was a generally warm interval of relatively high sea level, stratified oceans, and quasi-stable ecosystem conditions, but it was punctuated by brief episodes of abrupt change. Although most of the Cambrian has been characterized as a ‘greenhouse’ climatic state, the abruptness and synchroneity of sea level change strongly implies glacioeustasy.” PaleontologyIntroduction |
|||||||||||||||||||||||||
|
As a whole, the Cambrian metazoan fossil record is dominated numerically by arthropods, chiefly trilobites, of which there are over 100 Cambrian families. Much lower in abundance are Brachiopoda, represented chiefly by a number of inarticulate orders that may prove to merit class status. Hyoliths and certain members of the Mollusca, represented chiefly by monoplacophorans, form important though numerically minor elements.
“[S]tudies of Cambrian fossiliferous strata in Siberia, China, Europe, and Australia have yielded an unexpected range of well-preserved phosphatic microfossils. Most are either tiny shells or the disarticulated components (sclerites) of a protective armour comprised of many parts (Bengtson et al. 1990). These microfossils have become familiar as the ‘small shelly fossils.’ Most are original phosphatic skeletons or phosphatic copies of other kinds of hard parts ...” (Runnegar 1992 p. 66). Earliest Cambrian assemblages contain few metazoan taxa, though, and the diversity of trace and body fossils grew only over a protracted interval. However, the basal Cambrian may play host to more animal diversity than is sometimes recognised: Although reports of fossilised eggs and other early developmental stages of marine invertebrates are rare, probably due to the difficulties of recognizing them, there is an abundance of small globular structures in the fossil record, leading up to and including the earliest Cambrian. In 1994, Zhang and Pratt reported Middle Cambrian spherical fossils, 0.3 mm in diameter, that under a smooth membrane preserved a polygonal pattern which they interpreted as remains of blastomeres belonging to 64- and 128-cell stages of arthropod embryos. Shortly after, in 1997, Bengtson and Zhao reported similar occurrences of globular fossils from basal Cambrian rocks. Moreover, their finds included eggs containing identifiable embryos of metazoans (Fig. 4, Fig. 5) including a probable arthropod (also see below). |
(4)  9351 from the basal Cambrian Dengying Formation exposed in the Shizhonggou section, Shaanxi Province, China; scale is 500μm.") (5) (5)  32372, also from the Dengying Formation exposed in the Shizhonggou section. Length is about 1.8 mm.") Fig. 4: Reproduction of fig. 1A from Bengtson & Zhao 1997, a SEM image depicting a suggested metazoan embryo – possibly Olivooides multisulcatus – at approximately the 256-cell stage. Sample is NGMC (National Geological Museum of China) 9351 from the basal Cambrian Dengying Formation exposed in the Shizhonggou section, Shaanxi Province, China; scale is 500μm. Fig. 5: Reproduction of fig. 1K from Bengtson & Zhao 1997, a SEM image of the cnidarian Olivooides multisulcatus. Sample is CAGS (Chinese Academy of Geological Sciences) 32372, also from the Dengying Formation exposed in the Shizhonggou section. The length is about 1.8 mm. |
||||||||||||||||||||||||
Major Evolutionary EventsMajor TaxaThe most widely known of the ‘cast of characters’ from the Cambrian are probably the arthropods. The late Stephen Gould (1989) is largely responsible for the popularisation of the Cambrian Arthropoda, especially the problematic forms, since reinforced by the likes of Briggs et al. 1994 and Conway Morris 1998a, although anyone with the most casual interest in paleontology would have been aware of at least the Trilobita before the publication of any of those books. The Cambrian wasn’t all exoskeletons, however. In fact, definitive representatives of all readily fossilizable animal phyla (with the exception of bryozoans) have been found in Cambrian rocks, as have representatives of several soft-bodied phyla.Stromatolites are widely distributed in Cambrian carbonates, as are the calcified remains of photosynthetic organisms. Siliciclastic rocks yield abundant organic-walled microfossils, and many lithologies preserve a biogeochemical record of microbial diversity. Radiolaria, udoteacean green algae, and calcareous red algae may have evolved during the Cambrian, but they radiated during the Ordovician. (After Knoll 1996.) Sponges are generally accounted among the most primitive of Metazoans, although probable cnidarians pre-date them in the fossil record. Both were well-established long before the Cambrian and are known throughout the period. Sponges are a major component in the Early Cambrian Chengjiang and Qingjiang faunas, where they are second only to arthropods in both generic and specific diversity. Hundreds of sponge body fossils, distributed among more than 15 genera and twice as many species, have been collected there. “Archaeocyaths are an extinct group of sponges that had a very brief (geologically speaking) and spectacular history. The first archaeocyaths appear roughly 530 million years ago, during the Lower Cambrian. Archaeocyath species were very important members of Lower Cambrian communities. They diversified into hundreds of species during this time period and some of these species contributed greatly to the creation of the first reefs. Reef ecosystems tend to support a wide variety of organisms both in the present and in the past. Despite their great success in terms of numbers, the archaeocyaths were a short-lived group. They were almost completely non-existent by the middle Cambrian, some 10 to 15 million years after their first appearance” (Berkeley web site). Fossil ctenophores are seldom reported. Perhaps the earliest are the two Early Cambrian Chengjiang forms, Xianguangia sinica Chen & Erdtmann 1991 and Maotianoascus octonarius Chen & Zhou 1997, and an as-yet unnamed form from Qingjiang (see Fu et al. 2019, fig. 2C). From the Middle Cambrian comes Xanioascus canadensis Conway Morris & Collins 1996, known from the oldest of the Burgess Shale communities, found on Mt. Stephen in the Kicking Horse Shale Member. A slightly younger community from the Raymond Quarry Shale Member, includes Ctenorhabdotus capulus Conway Morris & Collins 1996. Probable cnidarians are known from Precambrian sediments of the Twitya Formation in the Mackenzie Mountains, Canada, dating from as early as 600 or 610 Ma. The Twitya fossils are simple cup-shaped animals, possibly similar to the sea anemones of today. Fasciculus vesanus Simonetta & Delle Cave 1978 is a possible cnidarian from the Burgess Shale. More complex forms controversially attributed to the Pennatulacea (sea pens) are known from the Ediacaran assemblages; if they are indeed pennatulaceans then they must have persisted through the Cambrian. Some other, more problematic, Ediacarans such as Kullingia also persisted into the Cambrian. “Hyoliths and halkierids (extinct forms thought to be related to mollusks), true conchiferan mollusks and, perhaps, chaetognaths enter the record during the first 10 to 12 million years of the Cambrian, but crown-group fossils of most other bilaterian phyla appear later: the earliest body fossils of brachiopods, arthropods, chordates, and echinoderms all post-date the beginning of the period by 10 to 25 million years” (Knoll & Carroll 1999). Gastropods, having planispiral shells with a wide, expanded aperture, make their first appearance in the Early Cambrian and seem to have been immediately and markedly successful. The earliest gastropods, such as Coreospira and Heliconella, are normally classified with the suborder Bellerophontacea although (a) they are not typical members of the suborder and (b) some authorities regard the Bellerophontacea as monoplacophorans rather than gastropods. Although the bivalves did not become common until Arenig (Ordovician) times, their minute, earliest representatives include Pojetaia, from the Early Cambrian of Australia (Runnegar & Bentley 1982) and Tuarangia, from the Middle Cambrian of New Zealand (McKinnon 1982). The former somewhat resembles the Ordovician palaeotaxodonts, and may have been an ancestor. Another Middle Cambrian form, Fordilla, is probably also a bivalve, though there is some debate on this point. (After Clarkson 1993, Ch. 8, pp. 191-253.) The fossil record of annelids is poor: Ediacaran forms such as Dickinsonia and Spriggina have been variously interpreted as annelids, but at least the latter has also been proposed as an arthropod and the best that can be said is that such interpretations are speculative. One of the earliest undoubted annelid fossils is the rare Early Cambrian Maotianchaeta fuxianella, known from Chengjiang. The evolution and phylogeny of the annelids is also poorly known; most authors now treat the oligochaetes as a (paraphyletic) stem group. |
|||||||||||||||||||||||||
|
Today the onychophorans are a special though almost insignificant arthropod sister-phylum represented by the terrestrial Peripatus and it’s allies. A number of Cambrian taxa have been referred to the Onychophora, however, despite being exclusively marine, and not without controversy. Microdictyon is one of the best known, and notable because its disarticulated plates are a common component of the so-called small shelly fauna.
Arthropods probably existed before the Cambrian; Ediacaran organisms such as Diplichnites and Parvancorina may have been arthropods. In the very basal Cambrian Dengying Formation (central Guizhou, China) the microfossil Markuelia secunda is thought to represent an arthropod embryo (Bengtson & Zhao 1997). By Chengjiang times, arthropods had diversified to include more than twenty five genera, including many of the exotica found in the much younger Burgess Shale, so thoroughly popularised by Gould 1989. By Burgess Shale times, stem-group arachnomorphs, such as the Nectaspida, had become both widespread and diverse, and, best known of all the extinct arthropods, the trilobites were also firmly established by this time, although their origins remain obscure. The earliest known brachiopods occur in Early Cambrian sediments and are already quite diverse. Three classes are recognised; and some authors consider the group to be polyphyletic. The Lingulata are a class of phosphatic-shelled inarticulate brachiopod; the genus Lingula arose in the Cambrian and persists today (possibly the best example of bradytelic evolution there is). The other classes, Articulata and Inarticulata, possess calcitic shells and are probably more closely related to each other than either is to the Lingulata. The earliest articulate group to evolve was the order Orthida. The earliest inarticulates may have appeared in the Middle Cambrian, but the group did not radiate substantially until the Middle Ordovician. |
(6) Fig. 6: Parvancorina minchami – A candidate Ediacaran arthropod, possibly a trilobite (see Fortey et al. 1996). In this scenario, the central axial ridge and the strongly arched anterior ‘lobes’ may be analogous to the midgut and gastric diverticulae. The scale bar is in centimetres. [Image and interpretation courtesy of Chris Nedin, Department of Industry, Science and Resources, Canberra.] Fig. 7: Redlichia (Hunanolenus) chinensis – Cambrian. Specimen from Henan Province, China. Length approximately 6 cm. [Original image.] |
||||||||||||||||||||||||
|
Echinoderms have a very extensive fossil record of about 13,000 species. Possible members (e.g. Arkarua) have been proposed from the Ediacaran fauna, but the group did not radiate strongly until the Cambrian. Cambrian representatives already possess ambulacral plates, indicating the presence of a water-vascular system and tube feet. Some of these early forms did not exhibit the pentaradial symmetry for which the group is now best known. The echinoderm classes living today, except the holothuroids, are known from the early Ordovician, while all other classes of the Cambrian radiation became extinct during the Paleozoic. (After Nielsen 2001, pp. 419-420.) The earliest pterobranchs, including the graptolites, first appear in the Middle Cambrian. The origins of graptolites are unknown (Rickards 1979) but the oldest fossil graptolites are the benthic dendroids (order Dendroidea; planktonic graptolites did not arise until earliest Ordovician, Tremadoc, time). These were colonial organisms having a complicated construction, and rather long stratigraphic ranges at the generic level. The dendroids persisted until the Carboniferous. Several genera of chordate were already established by the time of the Early Cambrian deposition at Chengjiang (Conway Morris 2000), including probable vertebrates such as Haikouichthys, Myllokunmingia, Zhongjianichthys, and Zhongxiniscusas. The more widely known Middle Cambrian Pikaia from the Burgess Shale may have been something of a laggard. “Conodonts were the first vertebrates to produce a mineralised skeleton and species of Protopanderodus and Periodon were some of the first animals to inhabit the deep-sea realm. Crown enamel in Protopanderodus and Periodon records seasonally entrained growth and cyclical alternation of episodes of growth and function. Growth episodes spanned 7-10 days in Protopanderodus graeai and only 2-3 days in Periodonaculeatus. It is postulated the former had a pelagic/nektonic mode of life whilst the latter was nektobenthic and specialised to living in deep-water habitats. The growth of P. graeai is further characterised by two distinct phases, the production of a triangular, symmetrical ‘proto-element’ followed by addition of increments of different thickness to the inner and outer edges. The curved and twisted geometry of the adult element was not initiated until the second phase of growth. This has implications for the function of the proto-element and the mode of life of juvenile animals” (Armstrong & Smith 1999). The earliest record of a possible land plant is the Cambrian age Aldanophyton Krishtofovich 1953, unfortunately known from only a single occurrence in Russia. The original, and perhaps only detailed description is in Russian; information in English about this enigmatic fossil is difficult to come by. It is treated as a junior synonym of the chlorophyte Margaretia Walcott 1931 by Conway Morris & Robison 1988. LagerstättenUnderstanding of Cambrian fossils, and their evolutionary significance, has been hugely enhanced by the study of fossils from several famous lagerstätten (sing. Lagerstätte): fossil localities which are highly remarkable for for either their diversity or quality of preservation; sometimes both.The most widely known of these is the Middle Cambrian Burgess Shale. Although this fossil site has been known since the early 1900s, very detailed study would not be accorded these fossils until the mid 1970s. The definitive popular account of this research, together with a contentious interpretation, is to be found in Stephen J. Gould’s book, Wonderful Life (1989). “In the 100 years since Walcott’s original discovery of the Burgess Shale, exceptionally preserved fossil assemblages have been reported from Cambrian strata of almost every paleocontinent” (Fu et al. 2019, p. 1338). The most important of these are significantly older than the Burgess Shale, though roughly contemporaneous with one another: the Chengjiang and Qingjiang localities in China, and the Sirius Passet locality in Greenland. Sirius Passet: Located on the northern coast of Greenland, this Early Cambrian site has yielded a diverse though not spectacularly preserved fauna, including a Redlichiid trilobite, Buenellus higginsi, the halkieriid Halkieria evangelista, and the enigmatic lobopod Kerygmachela kierkegaardi. The age is around 515 to 520 Ma. Chengjiang: Fossils from Chengjiang, near the city of Kunming in Yunnan Province, China, preserve a diverse biota of rather similar age to the Sirius Passet fauna; the differences between their fossil assemblages are attributed to environment and geography. The soft-bodied fossils include algae, medusiform metazoans, chondrophorines, sponges, chancelloriids, sea anemones, priapulid worms, hyoliths, possible ectoprocts, inarticulate brachiopods, annelid-like animals, lobopodians, trilobites and other arthropods, hemichordates, chordates as well as taxa that cannot definitely be assigned to any well established groups. Qingjiang: Qingjiang is a Burgess Shale/Chengjiang-type Cambrian lagerstätte, located in the Changyang area of South China. It is approximately contemporaneous with Chengjiang, but appears to have occupied a more distal environmental setting. Emu Bay Shale (Kangaroo Island): The Early Cambrian Emu Bay Shale of Kangaroo Island, South Australia, has long been famous as a source of magnificent specimens of the trilobites Redlichia takooensis and Hsuaspis bilobata. It is additionally important as the only site in Australia so far to yield a Burgess Shale-type biota. As such it represents an important basis for comparison with other Burgess Shale-type assemblages, particularly Chengjiang, which is the closest paleogeographically, although somewhat older. Burgess Shale: Middle Cambrian. I may come up with my own material eventually; meanwhile, do take a look at The Burgess Shale Geoscience foundation. ExtinctionsExtinction rates were generally high throughout the Cambrian, typically between 20 and 30%, but two major peaks in the mid Cambrian (Upper Solvan) and at the close of the Cambrian (Dolgellian) qualify as mass extinctions.Saltzman & Runkel (2001) note three significant paleoceanographic events occuring in Upper Cambrian sequences of Laurentia:
“The first two events are clearly global in scope, but the scale and timing of the sea level drop is not known in detail outside Laurentia. We have recently undertaken a project aimed at evaluating whether there are demonstrable cause and effect relationships among these events in widely separated Laurentian sections” (Saltzman & Runkel 2001). “The Cambrian–Ordovician represents a key interval in the evolution of the biosphere where several globally recognized extinctions of trilobite groups (i.e. ‘biomeres’) occurred (e.g. Saltzman et al. 2015). The causes of these extinctions are attributed to episodic upwelling of anoxic waters into shallow shelf environments during sea-level rise, an interpretation based on coeval positive d13C and d34S excursions inferred to reflect elevated organic and pyrite burial rates, respectively, under anoxic or euxinic conditions (Gill et al. 2011; Saltzman et al. 2015). If excursions of anoxic waters into shelf environments were the dominant driver of Cambrian–Ordovician trilobite extinctions, then both local and global proxies for anoxia are expected to provide complementary evidence as the cause of ecological turnover of marine communities and elevated extinction rates” (Edwards et al. 2018, p. 126). “There is a putative Late Cambrian lull in global phosphogenesis ... and SSF-style preservation that is hypothesized in the literature but not well demonstrated. If field testing demonstrates this lull to exist, then this period of decreased phosphogenesis could offer intriguing clues to potential oceanographic changes during the late Cambrian–earliest Ordovician lull in global diversity ... that preceded the Great Ordovician Biodiversification Event (GOBE…). “The diversity plateau that separates the Cambrian Explosion from the GOBE with its apparent temporary decrease in phosphogenesis ... provides potential clues to events that separate these two important times in the history of life on Earth. An understanding of fluctuations of the Cambrian-Ordovician phosphorus cycles may also inform understanding of the periodic extinction events ... that affected members of the Cambrian Fauna..., such as trilobites … and linguliform brachiopods.... “Recent studies have proposed ocean anoxia or euxinia as a factor in late Cambrian extinction patterns.... Anoxic bottom waters reduce the steepness of redox gradients and elevate the critical redox boundaries into the water column well above the sediment. This anoxia would lead to increased preservation of organic matter, the decay of which powers the Fe–P shuttle, ... and allow liberated phosphorus to diffuse into the water column.... Anoxia also inhibits sediment mixing by burrowing infauna, another source of redox oscillations ... and may thus reduce phosphate precipitation. Increased oceanic oxygenation has been identified as a factor that helped to end biomere extinctions..., and that triggered the onset of the GOBE.... This oxygenation would be predicted to coincide with increased phosphatic steinkern preservation, and this prediction does indeed seem to be the case” (Freeman et al. 2019). New Zealand OccurrencesFor most of the Paleozoic and Mesozoic, the rocks which would become the basement rocks of the Zealandia continent formed part of the Pacific margin of Gondwana, flanking Australia and Antarctica (Edbrooke 2017; Strogen et al. 2017). During this time, the Zealandia basement developed mainly by subduction-driven, episodic accretion. “Belts of Cambrian rocks with arc affinities in eastern Australia, Tasmania, New Zealand, and Antarctica are part of a single convergent margin, active over 30-40 Ma from the latest Early Cambrian to the Late Cambrian. … In the SE Australian and Tasmanian arc segments the proto-Gondwana plate subducted beneath the Pacific plate whereas subduction in the Antarctic and New Zealand segments was of opposite polarity. Common to most Cambrian fragments in SE proto-Gondwana is the tectonic overprint by the Ross-Delamerian orogeny from the Middle Cambrian to Early Ordovician…. A comparison of other Cambrian subduction-related rocks in Australia and Antarctica with the sections in New Zealand and Tasmania enables reconstruction of the nearly 5000-km-long Cambrian Pacific margin of SE proto-Gondwana. … At least one deformational event, possibly two, folded and disrupted the Cambrian sequence prior to the Devonian…. Both deformations indicate that a compressional tectonic setting in New Zealand had been present until the Late Cambrian” (Münker & Crawford 2000). In New Zealand, Cambrian rocks are known only from the Takaka Terrane, “the eastward of a pair of generally north-south trending terranes that form the ‘basement’ of west Nelson and parts of Fiordland” (Cooper 2004, p. 37; also see references therein). “Early Paleozoic rocks in New Zealand are confined to the western part of the South Island (Western Province) in Northwest Nelson, Westland, and Fiordland ... and the adjacent offshore region. Upper crustal sections of very low to low metamorphic grade are exposed in Northwest Nelson and Westland … whereas rocks exposed in Fiordland are their middle to lower crustal equivalents.... Cooper [1989] grouped early Paleozoic rocks of Northwest Nelson in two tectonostratigraphic terranes, namely, the Buller and Takaka Terranes.... Cambrian rocks are confined to the Takaka Terrane. This terrane comprises arc-related volcanic and volcaniclastic rocks, carbonates, cherts, and quartzo-feldspathic sediments in its Cambrian part. Conversely, a passive margin sequence of mainly nonvolcanic, quartz-rich clastic sediments and carbonates makes up the Ordovician-Silurian part of the Takaka T errane. Both the Buller and Takaka Terranes were amalgamated to the Gondwana margin at latest by the Devonian, when they were intruded by the Karamea Batholith” (Münker & Crawford 2000). ReferencesArmstrong, H.A.; Smith, C.J. 1999: Environmentally entrained growth in the earliest vertebrate teeth. Palaeontological Association 43rd Annual Meeting, University of Manchester, 19-22 December 1999 (Oral Presentation). Babcock, L.E.; Peng, S.C.; Brett, C.E.; Zhu, M.Y.; Ahlberg, P.; Bevis, M.; Robison, R.A. 2015: Global climate, sea level cycles, and biotic events in the Cambrian Period. Palaeoworld 24: 5-15. Bengtson, S.; Zhao, Y. 1997: Fossilized metazoan embryos from the earliest Cambrian. Science 277 (5332): 1645-1648. Bowring, S.A.; Erwin, D.H. 1998: A new look at evolutionary rates in deep time: Uniting paleontology and high-precision geochronology. GSA Today 8: 1-8. Briggs, D.E.G.; Erwin, Douglas H.; Collier, Frederick J.; Clark, C. 1994: The fossils of the Burgess Shale. Smithsonian Books: 1-256. Chen, J.; Erdtmann, B.-D. 1991: Lower Cambrian fossil lagerstätte from Chengjiang, Yunnan, china: Insights for reconstructing early metazoan life. In Simonetta, A.M.; Conway Morris, S. 1991: The early evolution of Metazoa and the significance of problematic taxa. Cambridge University Press: 57-76, pl. 1-3, text-fig. 1-7. Chen, J.; Zhou, G. 1997: Biology of the Chengjiang fauna. In Chen, J.; Cheng, Y.; Van Iten, H. (ed.) 1997: The Cambrian explosion and the fossil record. Bulletin of National Museum of Natural Science 10 10: 11-105. Clarkson, E.N.K. 1993: Invertebrate Paleontology and Evolution (third edition). Chapman and Hall. Cohen, K.M.; Finney, S.C.; Gibbard, P.L.; Fan, J.X. 2015: The ICS international chronostratigraphic chart v 2015/01. Episodes 36: 199-204. — 2017: The ICS international chronostratigraphic chart v 2017/02. Episodes 36: 199-204. Conway Morris, S. 1998a: The crucible of creation. Oxford University Press: 1-242. — 2000: The Cambrian "explosion": slow-fuse or megatonnage? Proceedings of the National Academy of Sciences of the USA 97 (9): 4426-4429. Conway Morris, S.; Collins, D.H. 1996: Middle Cambrian Ctenophores from the Stephen Formation, British Columbia, Canada. Ph Tr Roy Soc Lond 351: 279-308. Ph Tr RS Lond. Conway Morris, S.; Robison, R.A. 1988: More soft-bodied animals and algae from the Middle Cambrian of Utah and British Columbia. University of Kansas Paleontological Contributions 122: 1-48. Cooper, R.A. 1989: Early Paleozoic terranes of New Zealand. Journal of the Royal Society of New Zealand 19: 73-112. Cooper, R.A. (ed.) 2004: The New Zealand geologic timescale. Institute of Geological and Nuclear Sciences Monograph 22: 1-284. Edbrooke, S.W. 2017: The geological map of New Zealand. GNS Science Geological Map 2: 1-183. Edwards, C.T.; Fike, D.A.; Saltzman, M.R.; Lu, W.; Lu, Z. 2018: Evidence for local and global redox conditions at an Early Ordovician (Tremadocian) mass extinction. Earth and Planetary Science Letters 481: 125-135. Eichwald, C.E. 1840: Über das silurische Schichtensystems in Esthland. Zeitschrift für Natur-und Heilkunde der K. Medicinisch-chirurgische Akademie St. Petersburg 1/2: 1-210. Erwin, D.; Valentine, J. 2013: The Cambrian explosion: The construction of animal biodiversity. Freeman: 1-416. Fedonkin, M.A.; Gehling, J.G.; Grey, K.; Narbonne, G.M.; Vickers-Rich, P. 2007: The rise of animals: evolution and diversification of the Kingdom Animalia. Johns Hopkins: 1-326. Freeman, R.L.; Dattilo, B.F.; Brett, C.E. 2019: An integrated stratinomic model for the genesis and concentration of “small shelly fossil”-style phosphatic microsteinkerns in not-so-exceptional conditions. Palaeogeography, Palaeoclimatology, Palaeoecology 535: 109344. Fu, D.; Tong, G.; Dai, T.; Liu, W.; Yang, Y.; Zhang, Y.; Cui, L.; Li, L.; Yun, H.; Wu, Y.; Sun, A.; Liu, C.; Pei, W.; Gaines, R.R.; Zhang, X. 2019: The Qingjiang biota-A Burgess Shale-type fossil lagerstätte from the early Cambrian of South China. Science 363: 1338-1342. Gill, B.C.; Lyons, T.W.; Young, S.A.; Kump, L.R.; Knoll, A.H.; Saltzman, M.R. 2011: Geochemical evidence for widespread euxinia in the later Cambrian ocean. Nature 469: 80-83. Gould, S.J. 1989: Wonderful life. Penguin: 1-347. Gradstein, F.M.; Ogg, J.G.; Smith, A.G.; Bleeker, W.; Lourens, L.J. 2004: A new geologic time scale with special reference to Precambrian and Neogene. Episodes 27: 83-100. 27: 83-100. Harland, W.B.; Cox, A.V.; Llewellyn, P.G.; Tickton,C.A.; Smith, A.G.; Walters, R. 1982: A geological time scale. Cambridge University Press. Hearing, T.W.; Harvey, T.H.P.; Williams, M.; Leng, M.J.; Lamb, A.L.; Wilby, P.R.; Gabbott, S.E.; Pohl, A.; Donnadieu, Y. 2018: An early Cambrian greenhouse climate. Science Advances 4 (eaar5690): 1-11. Hou, X.; Aldridge, R.J.; Bergström, J.; Siveter, D.J.; Siveter, D.J.; Feng, X. 2004a: The Cambrian fossils of Chengjiang, China: The flowering of early animal life. Blackwell Publishing: 1-233. Knoll, A.H. 1996: Chapter 4. Archean and Proterozoic Paleontology. In Jansonius, J.; McGregor, D.C. (eds.) 1996: Paleontology: Principles and Applications. American Association of Stratigraphic Palynologists Foundation, v. 1, pp. 51-80. 1: 51-80. Knoll, A.H.; Carroll, S.B. 1999: Early animal evolution: Emerging views from comparative biology and geology. Science v. 284 (5423), issue of 25 Jun 1999, pp. 2129 - 2137. Science. Krishtofovich, A.N. 1953: Discovery of Lycopodiaceae in the Cambrian deposits of eastern Siberia [in Russian]. Doklady Akademii Nauk SSSR 91: 1377-1379. McKinnon, D.I. 1982: Tuarangia papura, n. gen. and n. sp. a late Middle Cambrian pelecypod from New Zealand. Journal of Paleontology 56: 589-598. Münker, C.; Crawford, A.J. 2000: Cambrian arc evolution along the SE Gondwana active margin: A synthesis ffrom Tasmania-New Zealand-Australia-Antarctica correlations. Tectonics 19 (3): 415-432. Nielsen, C. 2001: Animal evolution: Interrelationships of the living phyla (second edition). Oxford University Press: 1-378. Ogg, J.G.; Ogg, G.M.; Gradstein, F.M. 2016: A concise geologic time scale. Elsevier: 1-240. Rickards, R.B. 1979: Early evolution of graptolites and related groups. Systematics Association Special Volume 12: 435-442. Rogers, J.J.W. 1996: A history of the continents in the past three billion years. Journal of Geology 104: 91-107. Runnegar, B.; Bentley, C. 1982: Anatomy, ecology and affinities of the Australian Early Cambrian bivalve Pojetaia runnegari Jell. Journal of Paleontology 57: 73-92. Runnegar, B.N. 1992: Chapter 3 – Evolution of the Earliest Animals. In Schopf, J. W. (ed.) 1992a: Major Events in the History of Life. Jones and Bartlett, London: 65-93. Saltzman, M.R.; Edwards, C.T.; Adrain, J.M.; Westrop, S.R. 2015: Persistent oceanic anoxia and elevated extinction rates separate the Cambrian and Ordovician radiations. Geology 43: 807-810. Saltzman, M.R.; Runkel, A.C. 2001: Upper Cambrian extinction, carbon isotope shifts and the Sauk II - Sauk III Sea Level Event: new evidence from Northern Utah, Newfoundland and the midcontinent. North-Central Section - 35th Annual Meeting (April 23-24, 2001) Session No. 3 Paleozoic Bio-events, Illinois State University, Monday, April 23, 2001. Sedgwick, A.; Murchison, R.I. 1835: On the Silurian and Cambrian Systems, exhibiting the order in which the older sedimentary strata succeed each other in England and Wales. London and Edinburgh Philosophical Magazine and Journal of Science 7: 483-535. Seilacher, A. 1955: Spuren und fazies im Unterkambrium. In Schindewolf, O.; Seilacher, A. (ed.) 1955: Beiträge zur Kenntnis des Kambriums in der Salt Range (Pakistan). Akademie der Wissenschaften un der Literatur, Mainz, Abhandlungen der mathematisch-naturwissenschaftlichen Klasse 10: 261-446. Simonetta, A.; Delle Cave, L. 1978: Notes on new and strange Burgess Shale fossils (Middle Cambrian of British Columbia). Atti Soc. Tosc. Sci. Nat., Mem., Serie A 85: 45-49. Stirton, R.A. 1967: Time, Life, and Man. Wiley: 1-558. Strogen, D.P.; Seebeck, H.; Nicol, A.; King, P.R. 2017: Two-phase Cretaceous–Paleocene rifting in the Taranaki Basin region, New Zealand; implications for Gondwana breakup. Journal of the Geological Society, London 174: 929-946. van Eysinga, F.W.B. 1972: Geological time table (second edition). Elsevier. Walcott, C.D. 1931: Addenda to descriptions of Burgess shale fossils. Smithsonian Miscellaneous Collections 85 (3): 1-46. Wheeler, H.E. 1947: Base of the Cambrian System. Journal of Geology 55: 153-159. Zhang, X.; Pratt, B.R. 1994: Middle Cambrian arthropod embryos with blastomeres. Science 266: 627-639. Science. |
| Peripatus Home Page |
Hits counted from 1 May 2017:
My Traffic Estimate
 <i>pedum</i> defines the lower boundary of the Cambrian in the reference section at Fortune Head, southeastern Newfoundland. [Image courtesy of Dr. Gerd Geyer, Institut für Paläontologie, Bayerische Julius-Maximilians-Universität, Würzburg, Germany.]")
.")
. In this scenario, the central axial ridge and the strongly arched anterior ‘lobes’ may be analogous to the midgut and gastric diverticulae. The scale bar is in centimetres. [Image and interpretation courtesy of Chris Nedin, Department of Industry, Science and Resources, Canberra.]") (7)
(7)  <i>chinensis</i> – Cambrian. Specimen from Henan Province, China. Length approximately 6 cm. [Original image.]")