| Peripatus Home Page | Updated: 13-Feb-2024 |
AbstractThis page describes the Cretaceous Period, including stratigraphy, paleogeography, and famous lagerstðtten, followed by a sketched outline of some of the major evolutionary events.Keywords: Cretaceous, Cretaceous biota, fossil record, evolution IntroductionThe Cretaceous is the last period of the Mesozoic Era. It is probably most widely-known, at least popularly, for being the last period in which the dinosaurs held sway on land, pterosaurs in the air, and giant reptiles in the sea. But it was also the last stand of other smaller, though no less iconic animals from earlier times, notably the ammonites, belemnites, and rudist molluscs. There was no mass extinction event nor any great radiation event at the beginning of the Cretaceous; indeed, a convincing definition for Jurassic-Cretaceous boundary has been famously elusive. Animal and plant life appears to have crossed this boundary with little perturbation. Important evolutionary events occurred within the Cretaceous, however, perhaps most significantly the origin or at least the early radiation of the angiosperms (flowering plants), although conifer diversification was also important. Whether co-evolving with the angiosperms or on their own, insects were diversifying also, notably the Hymenoptera (ants and bees) but also aphids, butterflies and grasshoppers. The breakup of Pangea into separate continents continued, leading to increased plant and animal provincialism. The period was famously brought to a close by widespread volcanism centred on the Deccan plateau of southern India, and, at exactly the Cretaceous-Paleogene boundary (by definition), a bolide impact near Yucatan Peninsula. Both certainly contributed to one of the few greatest mass extinctions of the Phanerozoic at the end of the period. |
|
||||||||||
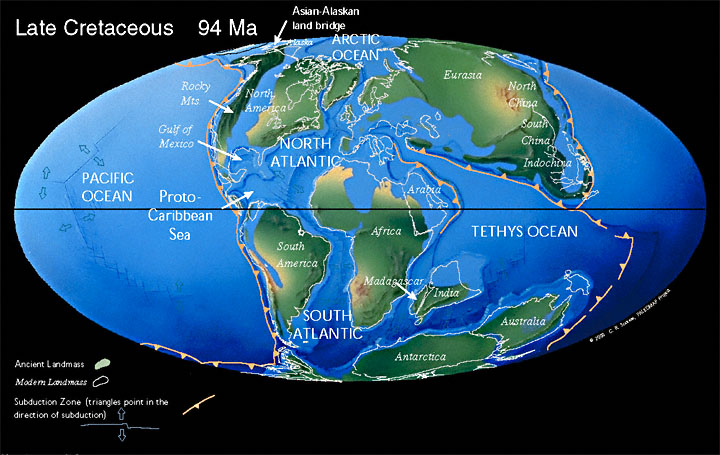
StratigraphyHistorical DevelopmentThe earliest concept of the Cretaceous was formailsed in 1822 when d’Halloy defined what he called the “Terraine Crétacé”. However, the base of the period has been modified over the years, most recently by the inclusion of the Berriasian as the lowermost stage in the Cretaceous System. Lower (Jurassic–Cretaceous) BoundaryThe base of the Cretaceous System is conventionally aligned with the base of the Berriasian Stage; that is, the Tithonian/Berriasian boundary is the Jurassic/Cretaceous boundary. As yet there is no ratified GSSP for the base of the Berriasian Stage although, by common usage, it lies near the first appearance datum (FAD) of the ammonite, Berriasella jacobi. Unfortunately, this ammonite is largely confined to the Mediterranean realm, so the datum is not much use internationally. At various times there have been proposals to redefine the Jurassic/Cretaceous boundary to align with the much more distinct Berriasian/Valanginian boundary (see Énay 2019 for an overview of the argument for such a move). These suggestions did not gain much traction initially, but may be gaining the upper hand, or at least serious consideration, more recently (Granier 2020). Upper (Cretaceous–Paleogene) BoundarySurely the most widely reported – and the most widely misreported – of all extinction events is that which brought the age of the dinosaurs to an end at the close of the Mesozoic: the “KT” extinction. Almost all of the popular and “lightly technical” literature published within the past couple of decades is filled with comet or meteorite impact theories, all ultimately traceable to the father and son team of Luis and Walter Alvarez. Indeed, there is an overwhelming body of evidence to support the contention that a large extraterrestrial impact did occur at the very end of the Cretaceous; so much so that the Cretaceous-Paleogene boundary is now defined by two impact proxies. The GSSP for the base of the Danian Stage (= Cretaceous-Paleogene boundary) was established in 1996 in the clay layer containing the famous “K/T” iridium anomaly and nickel-rich spinels at a section of Oued Djerfane, 8 km west of El Kef, in Tunisia (see Molina et al. 2006). ChronologyThe base of the Cretaceous, though not yet formally defined, is approximately 145 Ma. The top, however, has been very accurately dated: it is 66.04 Ma (Cohen et al. 2015, 2017). PaleogeographyMajor Tectonic EventsAt the beginning of the Cretaceous, Pangaea was breaking up into Laurasia in the north and Gondwana in the south. By the end of the period, at least the Gondwana continents had largely separated and were starting to migrate into their present day positions.During the Cretaceous, Zealandia continued to occupy a position on the proto-Pacific-facing, convergent margin of Gondwana, established since the Permian. Subduction on this margin ceased somewhere between 105 Ma and 70 Ma (approxmiately). The timing remains controversial, although Crampton et al. 2019 argues convincingly that subduction had finished on the New Zealand segment of the Gondwana margin by 100 Ma. This was followed by an extensional regime leading to the break-up of Gondwana, opening in the Tasman Sea, and thinning of the continental crust comprising Zealandia. Two temporally distinct phases of rifting have been recognized in the region: The first produced half-grabens trending NW to WNW during the mid-Cretaceous (c. 105 – 83 Ma). These rift basins are parallel to, but pre-date, the subsequent Tasman Sea spreading centres which were underway by approximately 82 Ma. They are followed by a short period (c. 83 – 80 Ma) of uplift and erosion, synchronous with Gondwana break-up, then a second rift phase during the latest Cretaceous–Paleocene (c. 80– 55 Ma) (abridged from Strogen et al. 2017). |
|||||||||||
Land and Sea“In the Mesozoic disintegration of eastern Gondwanaland, the Australo-Antarctic Gulf (AAG) formed as an eastward-propagating fracture between the future Antarctica and Australia. Floored by oceanic crust from about 100 million years ago (Ma), the AAG widened very slowly for about 50 myrs” (McGowran & Hill 2015, p. 20). |
(1) |
||||||||||
ClimatePaleontology
Major Taxa |
|||||||||||
Plant GroupsEarly Cretaceous vegetation was broadly similar to that of Late Jurassic times, both in distribution and general composition. Low paleolatitudes at that time were arid, having desert and sub-desert conditions, and here the floras were dominated by cheirolepidiacean conifers and matoniacean ferns. Northern mid paleolatitude floras were more diverse, including ferns, bennettitaleans, cycads, conifers and some ginkgos. At higher latitudes, diversity declined again, the floras dominated by leptostrobaleans and ginkgos. Southern mid paleolatitudes were dominated by bennettitaleans and cheirolepidiacean conifers. (Adapted from Cleal & Thomas 2009.) The most striking event in the evolution of plants during the Cretaceous was certainly the enormous radiation of angiosperms. The angiosperms (flowering plants) are the most diverse group of land plants living today, comprising some 270,000 described species – more than all other groups of land plants combined – placed in about 380 families and 83 orders (Mayr 2001, p. 64) and dominating modern plant ecosystems. “In theior rise to ecological dominance angiosperms have exhibited extraordinary developmental and evolutionary plasticity. This has resulted in overwhelming morphological diversity and a great variety of adaptive types. Angiosperms are far more diverse in vegetative form and in the structure of their reproductive organs than any other group of land plants” (Friis et al. 2011, p. 1). The group may have evolved from either the Gnetales or possibly the Bennettitales (Willis & McElwain 2002, p. 184). The enormous radiation of this group has largely occurred since the mid-Cretaceous, coevolving with a similar radiation of insects. However, the angiosperms must have arisen earlier: The most recent common ancestor of all living flowering plants is estimated to have existed perhaps as early as the Triassic or even the late Carboniferous (Qui et al. 1999) or, more conservatively, “between the Triassic and the Early Cretaceous (~247–136 million years ago” (Ramirez-Barahona et al. 2020). Evidence supporting the earliest date estimates is mainly provided by calibrated genetic divergence studies, though fossil angiosperm-like pollen and leaves have been found dating back to the late Triassic. Several form-species of Crinopolles-type pollen possessing a tectate wall have been described, dating to perhaps 220 Ma. The oldest leaves are somewhat younger, perhaps 210 Ma (Norian; Late Triassic), and include the problematic taxa Furcula and Sanmiguelia. A phylogenetic study by Silvestro et al. (2021), calibrated using fossil occurrence data, dates angiosperm origins in the Jurassic and the origin of angiosperm families such as the Arecaceae as mid Cretaceous. That being said, however, current orthodoxy is that the first true angiosperms evolved in the Early Cretaceous, probably in the Valanginian (~140 to ~133 Ma) or Hauterivian (~133 to ~129 Ma) ages. The oldest unequivocal angiosperm pollen grains first appear in the fossil record “during the Valanginian-Hauterivian; they spread out of the tropics in the Aptian and Albian [~125.0 to 100.5 Ma], and radiated in the Late Cretaceous” (Harris & Arens 2016, p. 640). Some of the earliest “body” fossils are “small plants, possibly rooted aquatics or wetland herbs” (Wing 2004, p. 90). “Compression flowers in general, however, are rare, and it is usually only the large ones which are seen. Some years ago, sieving techniques used for Tertiary sediments were applied to the Cretaceous, and yielded a previously unimagined diversity of Cretaceous angiosperm flowers, from sites in North America, Sweden, Portugal, Kazachstan, and Japan. It is now becoming clear how lineages are related. The earliest pollen is 135 million years old, and many basal eudicot lineages were fully established by about 110 Ma. Insect pollination is overwhelmingly supported by the evidence, and was probably important for enhancing speciation rates. Once started, the radiation of angiosperms, especially in low latitudes, kept rising, and shows no sign of levelling off in the Tertiary” (Clarkson 1999, p. 53). “The earliest unequivocal fossils assignable to angiosperms, probably representing small understorey plants thriving under warm climates, appear in the Early Cretaceous of northern Gondwana (~133–125 Ma), in areas roughly corresponding to the present-day Mediterranean region. Shortly thereafter, angiosperms experienced a major burst of morphological and ecological diversification, which by the middle Cretaceous (~115–100 Ma) had triggered the evolution of most extant lineages. Overwhelming palaeobotanical evidence indicates that, in many regions, the initial burst of diversification of angiosperms was not reflected in their ecological dominance” (Ramirez-Barahona et al. 2020; references elided). “Until the 1970s not a great deal was known about fossil flowers. Since then our knowledge has grown explosively. For example, the mid-Cretaceous Archaeanthus, from Russell, Kansas, now one of the best-known early flowers, has been the subject of extensive research. The flower is borne terminally on a long axis, and the seeds can be macerated out. The stamens and tepals are known from scars, resin bodies are scattered in the fruit and tepals, and all these features, together with the morphology, indicate an evident relation to the extant Magnoliacea” (Clarkson 1999, p. 53). Genetic evidence (Zanis et al. 2002) strongly suggests that the most ‘primitive’ (basal) living angiosperm is a little known shrub called Amborella trichopoda; a small shrub with tiny greenish-yellow flowers and red fruit, native to the South Pacific island of New Caledonia. The Nymphaeales (waterlilies and their relatives) are also contenders for the distinction of being the most basal living angiosperms. Perhaps the most basal fossil group yet to be well-delineated within the angiosperm clade is the Archaefructaceae, a family of herbaceous aquatic plants recovered from the Lower Cretaceous or possibly uppermost Jurassic Yixian Formation of western Liaoning, China. These plants had reproductive axes that lacked petals and sepals, and bore stamens in pairs below conduplicate (sharply folded together lengthwise) carpels. One combined morphological and molecular “total evidence” analysis places the Archaefructaceae as a sister group to all extant angiosperms, including Amborella and the Nymphaeales (Sun et al. 2002, p. 900). “As part of this mid- to Late Cretaceous angiosperm diversification, several key modern plant families appear early in the fossil record, including palms (Arecaceae),which are represented by leaf fossils in late Coniacian–early Santonian deposits from eastern USA and lower Campanian sites in Austria and North America” (Greenwood et al. 2022). |
(2) Fig. 2: Amborella trichopoda. [Photograph by Tim Stephens, courtesy of the Arboretum of the University of California, Santa Cruz.] |
||||||||||
BirdsBirds has already evolved by the latest Jurassic, at least, so the frequently repeated talk of “protofeathers” and other clues to the origin of birds being found in Cretaceous rocks (the Jehol literature is rife with this sort of thing) is patently ridiculous. Nevertheless, birds continued to evolve throughout the Cretaceous, when many important novelties appeared.The first known beak and pygostyle (the “parsons-nose” which is all that remains of the reptilian tail in modern birds) occur in a Chinese fossil dated at 130 Ma. Rahonavis is a primitive bird from 80 million-year-old rocks of Madagascar. Despite being more bird-like than Archaeopteryx, raven-sized Rahonavis retains some very distinctive theropod-like features. Other small primitive birds have been found elsewhere around the world. From Mongolia comes a large flightless bird, Mononykus, with wings replaced by a pair of single-digit hands that projected forwards. Another flightless bird is known from Patagonia. A sparrow-sized bird from Spain had a more modern shoulder joint than Archaeopteryx and a perching foot but it still had teeth. MammalsThroughout the early Mesozoic, mammals remained small, becoming more abundant, larger, and more diverse in the Cretaceous, which may have been a time of explosive radiation of Tribosphenida – early relatives of marsupials and placentals (Rougier 2002).“The diversity of living and extinct mammalian species is documented by the fossil record of ~220 million years and has evolved against the backdrop of radical alterations in terrestrial floras during the Cretaceous Terrestrial Revolution (KTR), the Cretaceous-Paleogene (KPg) mass extinction, continental rearrangements, and changes in key environmental parameters, such as average global temperature. However, the impact of these drivers on taxonomic diversification, particularly near the KPg boundary, remains controversial” (Meredith et al. 2011, p. 521). The results of the Meredith et al. (2011) study were “consistent with Benton’s hypothesis [Benton 2010] that the KTR (125 to 80 Ma), during which the angiosperm component of floras increased from 0 to 80%, was a key event in the diversification of mammals and birds” (p. 523). “[O]ur results are consistent with the hypothesis that both the KTR and the KPg mass extinction played important roles in the early diversification and adaptive radiation of mammals. The KTR increased ecospace diversity [the reality of ‘ecospace diversity’ appears to be an unconscious assumption present throughout this paper], possibly precipitating interordinal diversification, whereas the KPg mass extinction made more of this ecospace available for mammals, promoting the emergence of crown-group orders with their distinctive morphological adaptations” (Meredith et al. 2011, p. 523). LagerstðttenYixian Formation: Cretaceous (possibly some latest Jurassic? check this) Sihetun, Liaoning Province, China; true birds, dinosaurs, and several of the so-called ‘feathered dinosaurs’ [check if this is not the same as the previous entry]Jehol Group: Early Cretaceous Northeastern China; finds include the famous “feathered” dinosaurs, early birds, putative basal angiosperms, and primitive mammals. Detailed soft-tissue preservation of organisms is known. Las Hoyas: Located in Cuenca, Spain; Barremian in age. Most of the skeletal fossils appear articulated, and the exceptional preservation of them has allowed detailed study, obtaining information not available in other fossil sites. Hell Creek Formation: Montana and the Dakotas; non-marine fluvial channel-fill and floodplain deposits, primarily sandstone, siltstone and mudstone; diachronously overlain by lignites; almost entirely Late Cretaceous but in some places earliest Paleogene at the very top; iridium found in some of the overlying lignites may be related to the “K/T” iridium anomaly; dinosaurs have been known from the Hell Creek Formation since at least Barnum Brown’s AMNH expedition of 1902; diverse assemblage of theropods, ornithopods, pachycephalosaurs, ankylosaurs and ceratopsids, including the type and a few other specimens of Tyrannosaurus rex. See Lofgren 1997. Pierre Shale: Late Cretaceous (Campanian) North Dakota, USA; arthropods, vertebrates, including mosasaurs Hajoula Limestone: Cretaceous Lebanon; sublithographic limestone; fossil arthropods and fish Sierra de Montsech: Cretaceous Spain; fossil spiders, insects, crustaceans and vertebrates; see Selden 1989, 1990. Santana Formation: This spectacular locality is one of the most prolific sources of Early Cretaceous fish fossils. It is located in north east Brazil at the foot of Araripe Plateau, on the border of Ceará State. The fossils occur in shales, thin limestone bands, and commonly in rounded calcareous concretions. The site is most famous for fossil fish, but arthropods, molluscs, dinosaurs and pterosaurs, as well as some plants are also known. See Maisey 1991. ExtinctionsSurely the most widely reported – and in the lay media, at least, the most widely misreported – of all extinction events is that which brought the age of the dinosaurs to an end at the close of the Maastrichtian: the “KT” extinction. |
|||||||||||
|
The exact timing and nature of the end-Cretaceous mass-extinction event is famously contentious, so let us be clear about two things right from the outset: First, a bolide did strike the Earth at the end of the period. In fact, the Cretaceous-Paleogene boundary is essentially defined by this event [→ sidebar], so any suggestion that the impact occurred before or after the end of the Cretaceous is simply nonsensical. But, second, the almost unimaginably vast Deccan Traps volcanism was in full swing at the same time. Both of these events inevitably influenced the climate, the atmosphere, and life, to a great degree. One may reasonably argue about which event had the most influence over a particular group of organisms at a particular place and time, but to adopt one or other phenomenon as a complete explanation for the mass extinction, to the outright exclusion of the other, strikes me as ideological. That is not science.
“The idea of mass extinctions of life, traditionally by great floods, still has a strong hold on western imagination. Everyone has a favourite theory for major extinctions, all united by the common theme of attributing dominant importance to physical factors, and playing down the importance of normal biological mechanisms. Our own work strongly favours the diversification of both mammals and birds at least 30 million years before the extinction of dinosaurs – there must be ecological consequences for small dinosaurs from this early diversification” (Penny 2001). Fossil evidence also supports a progressive change in the composition of mammal communities across the K-T boundary, although dating uncertainties have complicated any simple interpretation of this data (e.g. Lofgren 1995). “Recent field and laboratory investigations have established that the latest Cretaceous (i.e. Campanian-Maastrichtian) sedimentary succession exposed within the James Ross Basin, Antarctica is in excess of 2 km in total thickness. Comprising essentially fine-grained, shallow-water, volcaniclastic rocks that are in places intensely fossiliferous, it represents one of the best opportunities to investigate palaeobiological and palaeoenvironmental changes leading up to the K-T boundary anywhere in the southern hemisphere. The exceptionally early extinction patterns of the inoceramid bivalves and belemnites can be confirmed, but it is apparent that other key groups such as the ammonites [also see Witts et al. 2018] and trigoniid bivalves go right up to the boundary itself. Studies throughout the 1000 m thick Maastrichtian sequence indicate that, although molluscan assemblages are abundant, they are never particularly diverse. The benthic element has a distinctly temperate aspect and there is both sedimentological and palaeontological evidence to suggest that it was subjected to periodic intervals of reduced oxygen levels. The comparatively small, but nevertheless still abrupt, extinction event at the end of the Cretaceous in Antarctica may well have been buffered to some extent by both the high-latitude position and unusual sedimentological setting of the basin” (Crame 1999). A number of authorities have reported multiple iridium spikes in the vicinity of the Cretaceous-Paleogene boundary (Ganapathy et al. 1981; Donovan et al. 1988; Graup & Spettel 1989; Bhandari et al. 1995, 1996; Zhao et al. 2002). These observations do not fit comfortably with the theory of a single, massive bolide impact being largely responsible for numerous end-Cretaceous phenomena. In the Nanxiong Basin, China, the evidence suggests that “the K/T event was not marked by an instantaneous geochemical environmental change, but stretched out over a considerable time” (Zhao et al. 2002, p. 10). Moreover, there is some evidence to support the view that the end-Cretaceous extinctions were more gradational, less severe, or both, at high paleolatitudes. For example, Crame et al. 1996 presents a study of Antarctic inoceramids and dimitobelids (a kind of belemnite), and concludes that “we cannot yet separate the effects of changes in water depths, water temperature and predation pressure with absolute certainty. We can, however, show that these groups became extinct exceptionally early in the Antarctic, and this fact alone enhances claims that the instability of polar water masses may have played an important role in bringing about the demise of at least some Late Cretaceous marine taxa” (p. 506). “Everyone has a favourite theory for major extinctions, all united by the common theme of attributing dominant importance to physical factors, and playing down the importance of normal biological mechanisms. Our own work strongly favours the diversification of both mammals and birds at least 30 million years before the extinction of dinosaurs – there must be ecological consequences for small dinosaurs from this early diversification” (Penny 2001). If an impact was the sole cause of the KT extinctions, then that would make the KT different from the several other mass extinctions known, which is not a parsimonious conclusion. However, an impact occurring at the same time time would certainly have added further stress to already failing ecosystems, and possibly accelerated or even heightened the extinction event in some places. |
|
||||||||||
New Zealand OccurrencesFor most of the Paleozoic and Mesozoic, the rocks which would become the basement rocks of the Zealandia continent formed part of the Pacific margin of Gondwana, flanking Australia and Antarctica (Edbrooke 2017; Strogen et al. 2017). During this time, the Zealandia basement developed mainly by subduction-driven, episodic accretion. Subduction at the Gondwana margin ended in late Early Cretaceous time, about 105 Ma. (Crampton et al. 2019 argues “that there is diverse evidence to indicate that subduction finished along the New Zealand segment of the Gondwana margin by 100 Ma.”) Following this, a period of extensional tectonics led to crustal thinning and the development of rift basins. By about 85 Ma, a margin-parallel rift basin opened completely, allowing inundation by what would become the Tasman Sea and Southwest Pacific Basin. Seafloor spreading and the production of new oceanic crust were well established by 80 Ma, as Zealandia drifted further away from Gondwana. (After Edbrooke 2017, p. 31.) ReferencesBenton, M.J. 2010: The origins of modern biodiversity on land. Philosophical Transactions of the Royal Society of London, series B: Biological Sciences 365: 3667-3679. Bhandari, A.; Shukla, P.N.; Ghevariya, Z.G.; Sundaram, S.M. 1995: Impact did not trigger Deccan volcanism: Evidence from Anjar K/T boundary layer in Deccan intertrappean sediments. Geophysics Research Letters 22: 433-436. Geo Res Lett. — 1996: K/T boundary layer in Deccan intertrappean at Anjar, Kutch. In Ryder, G. et al. (ed.) 1996: The Cretaceous-Tertiary event and other catastrophes in Earth history . Geological Society of America, Special Paper 307: 417-424. Clarkson, E.N.K. 1999: The Origin of Flowers - Association Annual Address. The Palaeontological Association Newsletter 41: 53. Cleal, C.J.; Thomas, B.A. 2009: An Introduction to Plant Fossils. Cambridge University Press: 1-248. Cohen, K.M.; Finney, S.C.; Gibbard, P.L.; Fan, J.X. 2015: The ICS international chronostratigraphic chart v 2015/01. Episodes 36: 199-204. — 2017: The ICS international chronostratigraphic chart v 2017/02. Episodes 36: 199-204. Crame, A. 1999: Changes in molluscan faunas across the K-T boundary in Antarctica. Palaeontological Association 43rd Annual Meeting, University of Manchester, 19-22 December 1999 (Oral Presentation). Crame, J.A.; Lomas, S.A.; Pirrie, D.; Luther, A. 1996: Late Cretaceous extinction patterns in Antarctica. Journal of the Gological Society, London 153: 503-506. Crampton, J.S.; Mortimer, N.; Bland, K.J.; Strogen, D.P.; Sagar, M.; Hines, B.R.; King, P.R.; Seebeck, H. 2019: Cretaceous termination of subduction at the Zealandia margin of Gondwana: The view from the paleo-trench. Gondwana Research 70: 222-242. dHalloy, J.G.J.dO. 1822: Observations sur un essai de cartes géologiques de la France, des Pays-Bas, et des contrées voisines. Annales de Mines 7: 353-376. Donovan, A.D.; Baum, G.R. et al. 1988: Sequence stratigraphic setting of the Cretaceous-Tertiary boundary in Central Alabama. In Wilgus, C.K. et al. (ed.) 1988: Sea-level changes - An integrated approach. Soc. Econ. Paleontol. Mineralog. Special Publication 42: 299-307. Edbrooke, S.W. 2017: The geological map of New Zealand. GNS Science Geological Map 2: 1-183. Énay, R. 2019: The Jurassic/Cretaceous system boundary is an impasse. Why do not go back to Oppel’s 1865 original an historic definition of the Tithonian? Cretaceous Research 106: 104241. Friis, E.M.; Crane, P.R.; Pedersen, K.R. 2011: Early flowers and angiosperm evolution. Cambridge University Press: 1-585. Ganapathy, R.; Gartner, S.; Jiang, M. 1981: Iridium anomaly at the Creataceous-Tertiary boundary in Texas. Earth and Planetary Science Letters 54: 393-396. E Pl Sci Lett. Gradstein, F.M.; Ogg, J.G.; Schmitz, M.D.; Ogg, G.M. 2012: The Geologic Time Scale 2012. Elsevier 1-2. Granier, B.R.C. 2020: Introduction to thematic issue, “The transition of the Jurassic to the Cretaceous: an early XXIth century holistic approach”. Cretaceous Research 114: 104530. Graup, G.; Spettel, B. 1989: Mineralogy and phase-chemistry of an Ir enriched pre-K/T layer from the Lattengebirge, Bavarian Alps, and significance for the KTB problem. Earth and Planetary Science Letters 95: 271-290. E Pl Sci Lett. Greenwood, D.R.; Conran, J.G.; K.West, C. 2022: Palm fronds from western Canada are the northernmost palms from the Late Cretaceous of North America and may include the oldest Arecaceae. Review of Palaeobotany and Palynology 301: e104641. Harris, E.B.; Arens, N.C. 2016: A mid-Cretaceous angiosperm-dominated macroflora from the Cedar Mountain Formation of Utah, USA. Journal of Paleontology 90 (4): 640-662. Lofgren, D.F. 1997: Hell Creek Formation. In Currie, P.J.; Padian, K. (ed.) 1997: Encyclopedia of dinosaurs. Academic Press: 302-303. Lofgren, D.L. 1995: The Bug Creek Problem and the Cretaceous-Tertiary Transition at McGuire Creek, Montana. University of California Press. Maisey, J.G. 1991: Santana fossils: an illustrated atlas. TFH Publications: 1-459. Mayr, E. 2001: What evolution is. Weidenfeld & Nicolson: 1-318. McGowran, B.; Hill, R.S. 2015: Cenozoic climatic shifts in southern Australia. Transactions of the Royal Society of South Australia 139 (1): 19-37. Meredith, R.W.; Janečka, J.E.; Gatesy, J.; Ryder, O.A.; Fisher, C.A.; Teeling, E.C.; Goodbla, A.; Eizirik, E.; Simão, T.L.L.; Stadler, T.; Rabosky, D.L.; Honeycutt, R.L.; Flynn, J.J.; Ingram, C.M.; Steiner, C.; Williams, T.L.; Robinson, T.J.; Burk-Herrick, A.; Westerman, M.; Ayoub, N.A.; Springer, M.S.; Murphy, W.J. 2011: Impacts of the Cretaceous terrestrial revolution and KPg extinction on mammal diversification. Science 334: 521-524. Molina, E.; Alegret, L.; Arenillas, I.; Arz, J.A.; Gallala, N.; Hardenbol, J.; von Salis, K.; Steurbaut, E.; Vandenberghe, N.; Zaghbib-Turki, D. 2006: The Global Boundary Stratotype Section and Point for the base of the Danian Stage (Paleocene, Paleogene, Tertiary, Cenozoic) at El Kef, Tunesia - Original definition and revision. Episodes 29: 263-273. Ogg, J.G.; Ogg, G.; Gradstein, F.M. 2008: The concise geologic time scale. Cambridge University Press: 1-177. Penny, D. 2001: Molecular Evolution: Introduction. Nature Encyclopedia of Life Sciences [doi:10.1038/npg.els.0001701]. Qui, Y.-L.; Lee, J.; Bernasconi-Quadroni, F.; Soltis, D.E.; Soltis, P.; Zanis, M.; Zimmer, E.A.; Chen, Z.; Savolainen, V.; Chase, M.W. 1999: The Earliest Angiosperms. Nature 402: 404-407. Ramírez-Barahona, S.; Sauquet, H.; Magallón, S. 2020: The delayed and geographically heterogeneous diversification of flowering plant families. Nature Ecology and Evolution 4: 1232-1238. Rougier, G.W. 2002: Mesozoic Mammals. In: eLS. John Wiley & Sons Ltd, Chichester. http://www.els.net [doi: 10.1038/npg.els.0001571]. Selden, P.A. 1989: Orb-weaving spiders in the early Cretaceous. Nature 340: 711-713. — 1990: Lower Cretaceous spiders from Sierra de Montsech, northeast Spain. Paleontology 33: 257-285. Silvestro, D.; Bacon, C.D.; Ding,W.; Zhang, Q.; Donoghue, P.C.J.; Antonelli, A.; Xing, Y. 2021: Fossil data support a pre-Cretaceous origin of flowering plants. Nat. Ecol. Evol. 5: 449-457. Strogen, D.P.; Seebeck, H.; Nicol, A.; King, P.R. 2017: Two-phase CretaceousPaleocene rifting in the Taranaki Basin region, New Zealand; implications for Gondwana breakup. Journal of the Geological Society, London 174: 929-946. Sun, G.; Ji, Q.; Dilcher, D.L.; Zheng, S.; Nixon, K.C.; Wang, X. 2002: Archaefructaceae, a new basal angiosperm family. Science 296: 899-904. Willis, K.J.; McElwain, J.C. 2002: The evolution of plants. Oxford: 1-378. Wing, S.L. 2004: Mass extinctions in plant evolution. In Taylor, P.D. (ed.) 2004: Extinctions in the history of life. Cambridge University Press: 61-97. Witts, J.D.; Landman, N.H.; Garb, M.P.; Boas, C.; Larina, E.; Rovelli, R.; Edwards, L.E.; Sherrell, R.M.; Cochran, J.K. 2018: A fossiliferous spherule-rich bed at the Cretaceous-Paleogene (K-Pg) boundary in Mississippi, USA: Implications for the K-Pg mass extinction event in the Mississippi Embayment and Eastern Gulf Coastal Plain. Cretaceous Research 91: 147-167. Zanis, M.J.; Soltis, D.E.; Soltis, P.S.; Mathews, S.; Donoghue, M.J. 2002: The root of the angiosperms revisited. Proceedings of the National Academy of Sciences of the USA 99: 6848-6853. Zhao, Z.; Mao, X.; Chai, Z.; Yang, G.; Kong, P.; Ebihara, M.; Zhao, Z. 2002: A possible causal relationship between the extinction of dinosaurs and K/T iridium enrichment in the Nanxiong Basin, South China: evidence from dinosaur eggshells. Palaeogeography, Palaeoclimatology, Palaeoecology 178: 1-17. |
| Peripatus Home Page |
Hits counted from 17 Aug 2018:
My Traffic Estimate