| Peripatus Home Page | Updated: 17-Sep-2024 |
AbstractThis page describes the Devonian Period, including stratigraphy, paleogeography, and famous lagerst‰tten, followed by a sketched outline of some of the major evolutionary events.Keywords: stratigraphy, Devonian Period, Devonian biota, fossil record, evolution, extinction, Rhynie Chert, Hunsruck Shale, Achanarras, Cleveland Shale, Canowindra, Escuminac Bay IntroductionBy the Devonian, life had become well-established on land: Scale tree forests began appearing, inhabited by the first wingless insects and, later, early amphibians. However, most Devonian biodiversity was still marine. Fishes were the dominant animals; blastoids are at their peak; the first ammonites evolved. |
|
||||||||||
StratigraphyType Section/SectionsThe Devonian was proposed by Adam Sedgwick and Roderick I. Murchison in 1839 (Sedgwick & Murchison 1839). The type section is in Devonshire; its boundaries are based mainly on fossils. Lower (Silurian-Devonian) BoundaryThe base of the Devonian is defined immediately at the first appearance of the graptolite species Monograptus uniformis in the rhythmically alterating limestones and calcareous shales of ‘Bed 20’ in the Klonk Section, which is located near the village of Suchomasty, about 35km southwest of Prague in the Czech Republic. The age is established at 419 ± 3.2 Ma. Upper (Devonian-Carboniferous) BoundaryThe GSSP defining the the base of the Carboniferous (i.e., the Devonian-Carboniferous Boundary) was established in 1991 (Paproth et al. 1991) at La Serre, France, where it is set at the first appearance datum (FAD) of the conodont Siphonodella sulcata. Subsequent research, however, has revealed stratigraphic problems with this datum, and it is now relised that this horizon cannot be correlated with precision at present. The GSSP is at the base of Bed 89 in Trench E at La Serre. The La Serre section is located in the southeastern Montagne Noire, Departement HÈrault, District of CabriËres, in southern France. Artificial trench E, which averages 80cm in depth on the southern slope of La Serre Hill, is about 125m south of the hilltop (252m), about 525m east of La Roquette farmhouse, 2.5km northeast of the village of FontËs (sheet 1:25,000 PÈzenas XXVI/44, 1-2, x = 682.55, y = 140.12). Sedimentology: The rocks in trench E’ are part of a geologic unit named “Klippen of CabriËres”. The youngest Devonian and earliest Carboniferous beds are characterized by a sequence of predominantly biodetrital oolitic limestone within a pelagic matrix of shale and cephalopod bearing calcilutites. Primary Markers: First appearance of the conodont Siphonodella sulcata within the evolutionary lineage from Siphonodella praesulcata to Siphonodella sulcata at the base of Bed 89 in trench E’. Secondary Markers: The trilobites Belgibole abruptirhachis, Archegonus (Phillibole) and Carbonocoryphe also occur in Bed 89. Some of these other taxa are especially important at other Locations around the world. For example, Belgibole abruptirhachis occurs immediately above the Hangenberg Schiefer interval in various cephalopod-bearing sections from the Renish Slate Mountains (Germany), the Holy Cross Mountains (Poland) and the Carnic Alps (Austria). See Paproth et al. 1991. ChronologyICS (Cohen et al. 2015) from 419.2 ± 3.2 to 358.9 ± 0.4 Ma. Paleogeography
|
|||||||||||
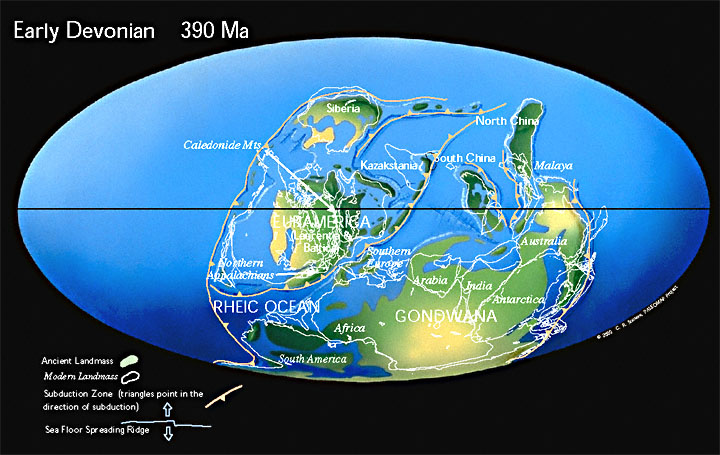
Land and SeaDuring the Devonian, there were three major continental masses: North America and Europe sat together near the equator, much of their current land underneath seas. To the north lay a portion of modern Siberia. A composite continent of South America, Africa, Antarctica, India, and Australia dominated the southern hemisphere. |
(1) |
||||||||||
|
PaleontologyIntroductionBy the Devonian, life had become well-established on land: Scale tree forests began appearing, inhabited by the first wingless insects and, later, early amphibians. However, most Devonian biodiversity was still marine. In the oceans, brachiopods flourished, along with tabulate and rugose corals, crinoids, and other echinoderms; blastoids are at their peak. Ammonites evolved and became common.However, the Devonian is probably most noted for the diversification of rise to prominence (and dominance) of various groups of fishes; indeed this period is occasionally referred to as the “age of fishes.” The terrestrial vegetation of the early Devonian consisted primarily of small plants, the tallest being only a meter tall. By the end of the Devonian, ferns, horsetails and seed plants had also appeared, producing the first trees and the first forests. Archaeopteris is one of these first trees. Early terrestrial arthropods, including wingless insects and the earliest arachnids. Tetrapods appear. Major Evolutionary Events“The Devonian marks a critical stage in the early evolution of vertebrates: It opens with an unprecedented diversity of fishes and closes with the earliest evidence of limbed tetrapods. However, the latter part of the Devonian has also been characterized as a period of global biotic crisis marked by two large extinction pulses: a “Big Five” mass extinction event at the Frasnian-Famennian stage boundary (374 Ma) [Kellwasser event] and the less well-documented Hangenberg event some 15 million years later at the Devonian-Carboniferous boundary (359 Ma)” (Sallan & Coates 2010, Abstract).Major TaxaPlants“From its primitive rhyniophyte and lycophyte precursors, land vegetation rapidly diversified during Devonian times. By the end of the period, all of the major divisions of vascular plants except the flowering plants had appeared, with the first record of ferns (or at least their pre-fern ancestors) being in Middle Devonian times, and of sphenophytes and gymnosperms in Late Devonian timesÖ” (Cleal & Thomas 2009, p. 203).“The Devonian was pivotal in the evolutionary history of land plants: a diverse range of plant architectures evolved and plants radiated into a remarkable array of ecological niches (Gensel & Andrews 1984, Gerrienne et al. 2004, Stein et al. 2012)” (Xu et al. 2017, p. 524). “The Devonian Period also saw the development of several structures that made the vascular plants better adapted to life on landÖ. Secondary wood is the most widely adopted means by which plants can significantly increase their stature and it is first seen in the fossil record in the Middle Devonian Series. The resulting trees (and therefore presumably forests) must have had a dramatic impact on the Devonian landscape. Photosynthetic efficiency was also enhanced by the development of planated leaves in Devonian times” (Cleal & Thomas 2009, p. 203). Numerous plant taxa have been reported from Devonian deposits around the world and phytogeographical provincialism began to develop in the global flora (see Raymond et al. 2006). However, plants with simple form and diminutive size, i.e., with axis widths less than 2 mm, from the Early–Middle Devonian, have been paid little attention (see Cai & Wang 1995, Wang & Berry 2003, Wang et al. 2004, 2007). Those basal euphyllophytes have quite distinct morphological characters in their branching patterns and sporangial attachment, and played key roles in the evolution of early land plants (Stewart & Rothwell 1993, Kenrick & Crane 1997)” (Xu et al. 2017, p. 524; Fig. 2). The lycopsids apparently arose in the Silurian (Kotyk et al. 2002) but their fossil record is better known from the Devonian onwards. The sphenophytes appear to have arisen in the Devonian. |
(2) Fig. 2: The Early Devonian plant, Pauthecophyton; composite from Xu et al. 2019, fig. 3A and D. |
||||||||||
|
Bryophyta higher plants Mollusca Annelida Arthropoda
Brachiopoda Echinodermata graptolites FishVertebrates underwent a major adaptive radiation during the Devonian, “when jawed species (gnathostomes) and particularly placoderms (armoured fishes), became dominant. A Lochkovian [earliest Devonian] peak of diversity is registered in various Lower Devonian series all around the Old Red Sandstone Continent and Siberia, for ostracoderms in general, and heterostracan pteraspidomorphs in particular. It occurs at different time slices in the Lochkovian, depending on the localities, and may be followed by another smaller peak in the Pragian” (Blieck 2017, Abstract).“The rise of jawed vertebrates (gnathostomes) throughout the Devonian ... and into the post-Devonian is one of the key episodes in vertebrate evolution.... This interval encompasses well-known early diversification events, including those of Osteichthyes (bony fishes: ray-finned Actinopterygii and lobe-finned Sarcopterygii, including tetrapods), Chondrichthyes (cartilaginous fishes: Elasmobranchii and Holocephalii), and Placodermi and Acanthodii (extinct groups of debated affinity to extant gnathostomes)” (Sallan & Coates 2010, p. 10131). Placoderms were an extinct group of jawed fish, first appearing in the Late Silurian Period and rising to dominance in the Devonian Period. A few straggled on into the Early Carboniferous (Mississippian), but mostly they had done their dash by then. These were armoured fish: the front half of their bodies was covered in large bony plates, forming a rigid shield with a joint between the head and shoulder. There were two major groups of placoderms: the antiarchs and the arthrodires. The arthrodires had a ball-and-socket joint between the shoulder and head which permitted the head to be rotated back against the body, providing the mouth with a wide gape. Most arthrodires had large eyes and powerful jaws lined with sharp-edged plates of bone, and they were undoubtedly predators. The remainder of the body seems to have lacked scales; the only bony traces found have been vertebrae and rays supporting the medial fins. The structure of the pectoral fins is unknown, but large sockets on each side of the shoulder shield suggest great mobility and arthrodires are assumed to have been fast-moving, agile swimmers. The earliest known sharks are Silurian, and the group was highly diversified by the Devonian. Some of the Devonian sharks, such as the 1.8 m Cladoselache, were certainly apex predators, but perhaps the best known apex predator from this period was the arthrodire placoderm, Dunkleosteus. This animal is usually depicted with long ribbon-like fins top and bottom, but in fact the post-cranial anatomy is poorly known from relatively few fossils. There is certainly quite a range of opinions about its size presented on the internet, many of them quite far-fetched. Generally, the more reputable sources give the lower estimates. The Australian vertebrate paleontologist, John Long, gives an estimate of 6 metres in his 1995 book on fossil fishes, and the AMNH web site says the same (“20 feet”). To put that into some kind of context, this is bigger than a marlin, about the same size as the very largest great white sharks of today, and slightly smaller than a large male Orca. The oldest lobe-fin fossil fish are Early Devonian, though the group likely appeared earlier. tetrapods – see Clack 2002a TetrapodsTetrapodomorphs comprise “the limbed tetrapods and their closest fish relatives, whose earliest record is from the Pragian of China (Lu et al. 2012). The group diversified greatly in both marine and freshwater habitats during the Middle-to-Late Devonian while giving rise to several distinct lineages, including the earliest limbed tetrapods (Ahlberg 2018). Whereas the tetrapods flourished after the Devonian, limbless fish-grade tetrapodomorphs underwent a marked reduction in diversity during the Carboniferous, with only a handful of representatives persisting into the early Permian before vanishing from the fossil record (Romano et al. 2014)” (Choo et al. 2024).Zhu & Yu (2002) describe a new fossil sarcopterygian fish, Styloichthys changae, that “possesses an eyestalk and which exhibits the character combination expected in a stem group close to the last common ancestor of tetrapods and lungfish. Styloichthys from the Lower Devonian of China bridges the morphological gap between stem-group sarcopterygians (Psarolepis and Achoania) and basal tetrapodomorphs/basal dipnomorphs. It provides information that will help in the study of the relationship of early sarcopterygians, and which will also help to resolve the tetrapod-lungfish divergence into a documented sequence of character acquisition” (Zhu & Yu 2002, p. 767, Abstract). The gnathostomes (jawed vertebrates) “underwent major changes over the Devonian-Mississippian divide.... Placoderms, sarcopterygians, and acanthodians are replaced by chondrichthyans, actinopterygians, and tetrapods, occupying a wider range of ecological roles and dominating all succeeding biotas.... This faunal transformation has been subjected to few analyses, and explanations have tended to focus on gradual replacement ... and competitive displacementÖ” (Sallan & Coates 2010, p. 10131). “Most of our understanding about these crucial vertebrate events comes from only a few freshwater deposits from Euramerican localities, with outcrops in Greenland producing the most prolific stegocephalian remains. Elginerpeton and Obruchevichthys were first, between 385 and 375 MYBP, followed several million years later by a modest radiation that included Ventastega, Ichythostega, Acanthostega, Tulerpeton, Metaxygnathus, and Hynerpeton. Although most of these taxa possessed limbs (although this is not certain for Elginerpeton and Obruchevichthys), all of these taxa have been interpreted as fully aquatic, rather than terrestrial.... Their respiratory systems are poorly known, but osteology suggests that they were able to derive some amount of O2 from water instead of being entirely air breathing.... These stegocephalians disappeared soon thereafter, followed rapidly by rare appearances of limbed vertebrates” (Ward et al. 2006; citations omitted). Lagerst‰ttenSome of the several famous lagerstätten of Devonian age are briefly described below.Rhynie Chert: The Rhynie Chert in Scotland is an Early Devonian age deposit containing fossils of both Zosterophyllophytes and Trimerophytes, the two major lines of vascular plants. This indicates that prior to the start of the Devonian, the first major radiations of the plants had already happened. It has become famous as the oldest known terrestrial ecosystem fossilised in place. Hunsrück Schiefer (Hunsrück Slate): The Hunsr¸ck Slate formation, originally and still mined for roofing slates, is one of the most important Devonian fossil localities known. Although the fossils are generally not spectacular in a manner which appeals to amateurs, the unusual preservation of soft tissue by pyritisation makes these rock a unique archive of scientific information. The rock is a marine sediment preserving the usual arthropods and molluscs, but also plants, sponges, vertebrates, and even extremely delicate forms such as ctenophores and chondrophorans. Gilboa: M. Dev; New York State; spiders and pseudoscorpions; Shear et al. 1984, Selden et al. 1991 Achanarras Fauna (Scotland): “The classic Tynet Burn and Gamrie fish localities, in north-east Scotland are of Middle Devonian (Eifelian) age and have, since last century yielded beautifully preserved fossil fish, typical of the Achanarras fauna which occurs in deposits exposed to the south of the Moray Firth and to the far North. The fish are mostly preserved in calcareous concretions and despite the large quantities of specimens that have come from the sites, no special significance has been attributed to the material with respect to unnusual preservation. Current detailed work on newly collected material however, has identified new preservational potential for the fish bearing concretions. A number of rare specimens of acanthodians from both sites consistently exhibit dark traces within the body cavity which are analogous with the sites of internal organs in modern groups. Furthermore, dark ‘spots’ in the heads of acanthodians from Gamrie can be interpreted as the position of the eye. To date, these structures have only been observed in three acanthodian genera, but other groups are present and clearly there is potential for more work. A museum collection survey has been initiated and two more specimens, collected last century, have been located which exhibit internal organ site preservation” (Davidson & Trewin 1999). Cleveland Shale: Late Devonian (Fammenian); found near Cleveland, Ohio, USA; a vertebrate lagerstätte containing articulated specimens of the cladodont sharks Cladoselache (several species), Ctenacanthus compressus, and the coronodontid shark Diademodus hydei (the holotype and only specimen of this shark); although some fossils occur in the shale itself, most occur in flattened discoidal dolomitic concretions which preserve soft tissues, such as muscle fibers, outlines of the dermal membrane of the body and fins, and ingested prey. Two Cladoselache specimens, one of C. fyleri and one of C. kepleri, also have preserved kidneys. Two specimens of the arthrodire Dunkleosteus terrelli, an articulated specimen from a concretion and one in cone-in-cone, also have a pectoral fin partially preserved. (Thanks to Douglas Dunn, Cleveland Museum of Natural History, for the detailed information.) Canowindra: Late Devonian; famous fish locality Escuminac Bay: Late Devonian (early Frasnian); eastern Canada, near the village of Miguasha; anaspids, placoderms, and other fishes including Eusthenopteron; Schultz & Cloutier 1996, Clack 2002a (p. 87) Gogo Formation: Late Devonian (early Frasnian); Western Australia; 50 species of fishes, concavicarid crustaceans, rare eurypterids, tentaculatids and other invertebrates; Long & Trinajstic 2010, Long & Trinajstic 2018 ExtinctionsTwo major mass extinctions occurred late in the Devonian: the Kellwasser event at the Frasnian-Famennian stage boundary, and the Hangenberg event at the end of the period, at the Devonian-Carboniferous boundary.Kellwasser eventThe Kellwasser event of the Frasnian-Famennian stage boundary “is associated with spectacular losses in marine diversity involving ~13–40% of families and ~50–60% of genera (McGhee 1996, Caplan & Bustin 1999)” (Sallan & Coates 2010, p. 10131). “The Frasnian-Famennian (F-F) faunal crisis represents one of the ‘Big Five Mass Extinctions’ in Earth History. Studies have been published that favor either a collision with an extraterrestrial bolide ... or changes in the oceanographic and climatic systems .... However, no unequivocal evidence on the origin of the Late Devonian faunal breakdown was reported. E.g. iridium anomalies reported from Australia ... and Southern China ... post-date the F-F boundary by 1.5 to 2 Ma. Microtekites from Belgium boundary sections ... were seen in context with the 368±1 Ma old Siljan Ring impact structure. However, the cosmic origin of these spherules has been questioned ... and the microtektites were found above the F-F boundary and clearly post-date the extinction event. Negative carbon isotope excursions were interpreted as evidence for a sudden biomass crash hat may have been triggered by a bolide impact .... In contrast, two positive δ13C excursions measured in the late Frasnian and at the F-F boundary ... seem not to support the idea of a dramatic decline in primary productivity. “The geochemical data base for the Late Devonian extinction event shows no evidence for a bolide impact as potential cause of the mass extinction. Instead, repeated changes in the carbon cycle of the ocean-atmosphere system are indicated by positive carbon excursions. The enhanced organic carbon burial is indicated by the higher δ13C values, deposition of the Kellwasser horizon and a positive excursion in δ34S of sulfides and organically-bound sulfur A decrease in atmospheric and oceanic dissolved CO2 contents is expected and may have culminated in global climatic cooling. A decrease in tropical seasurface temperature of 7°C is indicated by preliminary conodont and fish apatite δ18O data. “The paleontological data base seems to support the conclusion that climatic cooling may have represented a potential mechanism for the Late Devonian mass extinction .... Organisms living in the tropical to subtropical pelagic and shallow-water ecosystems were heavily decimated. Organisms thriving in higher latitudes or in deeper waters were only slightly affected. Further, late Frasnian faunal groups that were adapted to cooler temperatures migrated into tropical latitudes during the early Famennian. This pattern suggests that climatic cooling in conjunction with significant oceanographic changes may represent a powerful scenario to account for the Late Devonian mass extinction” (Joachimski & Buggisch 2000; ellipses replace numbered references in the original). Hangenberg eventFollowing on the heels of the Late Devonian Kellwasser extinction event, the end-Devonian mass extinction, or Hangenberg Crisis, at 359 Ma (Myrow et al. 2013), is now also considered among the first-order mass extinctions. “The extinction was a multiple biotic crisis with an early main phase of marine extinctions associated with black shales and anoxia (Kaiser et al. 2016) separated by an estimated 300 thousand years (ka) (Myrow et al. 2013) from a terrestrial mass extinction of fish (Sallan & Coates 2010) with coincident extinctions (Silvestro et al. 2015) in land plants. Regarding pollen and spores, the terrestrial extinction is clearly expressed as the complete loss of diversity (Higgs et al. 1988, van Veen 1981) across the Devonian-Carboniferous (D-C) boundary with the extinction of at least four major spore groups that had dominated the spore assemblage (van Veen 1981). The immediate post-extinction VI spore assemblage was remarkable for its simplicity (Higgs et al. 1988) with the recovery vegetation dominated by small simple land plants. Hence, the event was a major ecosystem disruption that included the collapse of the structured forest community (Greb et al. 2006)” (Marshall et al. 2020, p. 1). Sallan & Coates (2010) notes that “major vertebrate clades suffered acute and systematic effects centered on the Hangenberg extinction involving long-term losses of over 50% of diversity and the restructuring of vertebrate ecosystems worldwide. Marine and nonmarine faunas were equally affected, precluding the existence of environmental refugia. The subsequent recovery of previously diverse groups (including placoderms, sarcopterygian fish, and acanthodians) was minimal. Tetrapods, actinopterygians, and chondrichthyans, all scarce within the Devonian, undergo large diversification events in the aftermath of the extinction, dominating all subsequent faunas. The Hangenberg event represents a previously unrecognized bottleneck in the evolutionary history of vertebrates as a whole and a historical contingency that shaped the roots of modern biodiversity” (Sallan & Coates 2010, Abstract). “Every ecological ordination analysis applied to the total matrix [cluster analysis, canonical correspondence analysis..., nonparametric multidimensional scaling..., and factor analysis...] shows the same basic pattern.... Devonian faunas are distinct from Mississippian faunas.... [W]e conclude that the end-Devonian Hangenberg event was a first-order magnitude extinction for jawed vertebrates comparable to Big Five events, including the end-Cretaceous and end-Permian.... Furthermore, the Hangenberg extinction was a global phenomenon, affecting all ecosystems no matter how the data are analyzed” (Sallan & Coates 2010, p. 10132, 10134). Marshall et al. 2020 advance the idea that the extinction was due to ozone layer depletion during a period of rapid warming, caused (citing Anderson et al. 2012) by increased convective transport of water vapour into the stratosphere. As evidence they present malformed spores and spore tetrads, claiming these are “entirely consistent Ö with exposure to elevated levels of ultraviolet-B (UV-B) that damaged the DNA before the formation of their protective sporopollenin walls” indicating “the temporary loss of the global protective ozone layer.” While intriguing, this hypothesis is rather weakly supported to date, and other scenarios are equally viable. New Zealand OccurrencesFor most of the Paleozoic and Mesozoic, the rocks which would become the basement rocks of the Zealandia continent formed part of the Pacific margin of Gondwana, flanking Australia and Antarctica (Edbrooke 2017; Strogen et al. 2017). During this time, the Zealandia basement developed mainly by subduction-driven, episodic accretion. Devonian rocks are rare in New Zealand. The best known and most accessible are to be found near Reefton, at Lankey Creek and Stoney Creek, where fossil brachiopods are relatively common. Some other animals, such as corals, also occur but they are less common. Very rare trilobites are also known. ReferencesAhlberg, P.E. 2018: Follow the footprints and mind the gaps: a new look at the origin of tetrapods. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 109 (1-2): 115-137. Anderson, J.G.; Wilmouth, D.M.; Smith, J.B.; Sayres, D.S. 2012: UV dosage levels in summer: Increased risk of ozone loss from convectively injected water vapour. Science 337: 835-839. Blieck, A. 2017: Heterostracan vertebrates and the Great Eodevonian Biodiversification Event — an essay. Palaeobiodiversity and Palaeoenvironments 97 (3): 375-390. Cai, C.; Wang, Y. 1995: Devonian floras. In Li, X. (ed.) 1995: Fossil floras of China through the geological ages. Guangdong Science and Technology Press, Guangzhou: 28-77. Caplan, M.L.; Bustin, R.M. 1999: Devonian-Carboniferous mass extinction event, widespread organic-rich mudrock and anoxia: Causes and consequences. Palaeogeography, Palaeoclimatology, Palaeoecology 148: 187-207. Choo, B.; Holland, T.; Clement, A.M.; King, B.; Challands, T.; Young, G.; Long, J.A. 2024: A new stem-tetrapod fish from the Middle-Late Devonian of central Australia. Journal of Vertebrate Paleontology: 1-15. Clack, J.A. 2002: Gaining ground: The Origin and Early Evolution of Tetrapods. Indiana University Press: 1-400. Cleal, C.J.; Thomas, B.A. 2009: An Introduction to Plant Fossils. Cambridge University Press: 1-248. Cohen, K.M.; Finney, S.C.; Gibbard, P.L.; Fan, J.X. 2015: The ICS international chronostratigraphic chart v 2015/01. Episodes 36: 199-204. Davidson, R.G.; Trewin, N.H. 1999: Unusual Soft Tissue Preservation in Middle Devonian Fish-Bearing Nodule Beds. Palaeontological Association 43rd Annual Meeting, University of Manchester, 19-22 December 1999. Edbrooke, S.W. 2017: The geological map of New Zealand. GNS Science Geological Map 2: 1-183. Gensel, P.G.; Andrews, H.N. 1984: Plant Life in the Devonian. Praeger, New York: 1-381. Gerrienne, P.; Meyer-Berthaud, B.; Fairon-Demaret, M.; Streel, M.; Steemans, P. 2004: Runcaria, a Middle Devonian seed plant precursor. Science 306: 856-858. Greb, S.F.; DiMichele, W.A.; Gastaldo, R.A. 2006: Evolution and importance of wetlands in earthhistory. Geol. Soc. Am. Spec. Pap. 399: 1-40. Higgs, K.; Clayton, G.; Keegan, J.B. 1988: Stratigraphic and systematic palynology of theTournaisian Rocks of Ireland. Geol. Surv. Ireland Spec. Pap. 7: 1-93. Kaiser, S.I.; Aretz, M.; Becker, R.T. 2016: The global Hangenberg Crisis (Devonian-Carboniferoustransition): Review of a first-order mass extinction. Geol. Soc. Lond. Spec. Pub. 423: 387-437. Kenrick, P.; Crane, P.R. 1997: The origin and early evolution of plants on land. Nature 389: 33-39. Kotyk, M.E.; Basinger, J.F.; Gensel, P.G.; de Freitas, T.A. 2002: Morphologically complex plant macrofossils from the late Silurian of Arctic Canada. American Journal of Botany 89 (6): 1004-1013. Long, J.A.; Trinajstic, K.M. 2010: The Late Devonian Gogo Formation Lagerstätte - Exceptional preservation and diversity in early vertebrates. Annual Reviews of Earth and Planetary Sciences 38: 665-680. — 2018: A review of recent discoveries of exceptionally preserved fossil fishes from the Gogo sites (Late Devonian, Western Australia). Earth and Environmental Science Transactions of the Royal Society of Edinburgh 108: 111-117. Lu, J.; Zhu, M.; Long, J.A.; Zhao, W.; Senden, T.J.; Jia L.T.; Qiao, T. 2012: . The earliest known stem-tetrapod from the Lower Devonian of China. Nature Communications 3: 1160. Marshall, J.E.A.; Lakin, J.; Troth, I.; Wallace-Johnson, S.M. 2020: UV-B radiation was the Devonian-Carboniferous boundary terrestrial extinction kill mechanism. Science Advances 6 (eaba0768): 1-8. McGhee, G.R. 1996: The Late Devonian Mass Extinction: The Frasnian-Famennian Crisis. Columbia University Press. Myrow, P.M.; Ramezani, J.; Hanson, A.E.; Bowring, S.A.; Racki, G.; Rakociński, M. 2013: High-precision U-Pb age and duration of the latest Devonian (Famennian) Hangenberg event, and its implications. Terra Nova 26: 222-229. Ogg, J.G.; Ogg, G.; Gradstein, F.M. 2008: The concise geologic time scale. Cambridge University Press: 1-177. Paproth, E., Feist, R., and Flaijs, G. 1991: Decision on the Devonian-Carboniferous boundary stratotype. Episodes 14 (4): 331-336. Raymond, A.; Gensel, P.G.; Stein, W.E. 2006: Phytogeography of late Silurian macrofloras. Review of Palaeobotany and Palynology 142: 165-192. Romano, C.; Koot, M.A.; Kogan, I.; Brayard, A.; Minikh, A.V.; Brinkmann, W.; Bucher, H.; Kriwet, J. 2014: Permian-Triassic Osteichthyes (bony fishes): diversity dynamics and body size evolution. Biological Reviews 91: 106-147. Sallan, L.C.; Coates, M.I. 2010: End-Devonian extinction and a bottleneck in the early evolution of modern jawed vertebrates. Proceedings of the National Academy of Sciences of the USA 107 (22): 10131-10135. Schultze, H.-P.; Cloutier, R. (ed) 1996: Devonian fishes and plants of Miguasha, Quebec, Canada. Verlag Dr. Friedrich Pfeil, Munchen: 1-374. Sedgwick, A.; Murchison, R.I. 1839: On the physical structure of Devonshire, and on the subdivisions and geological relations of its older stratified deposits. Transactions of the Geological Society of London, Series 2, no. 5: 633-704. Selden, P.A.; Shear, W.A.; Bonamo, P.M. 1991: A spider and other arachnids from the Devonian of New York, and reinterpretations of Devonian Araneae. Paleontology 34 (2): 241-281. Shear, W.A.; Bonamo, P.M.; Grierson, J.D.; Rolfe, W.D.I.; Smith, E.L.; Norton, R.A. 1984: Early land animals in North America: evidence from Devonian age arthropods from Gilboa, New York. Science 224 (4648): 492-494. Silvestro, D.; Cascales-Miñana, B.; Bacon, C.D.; Antonelli, A. 2015: Revisiting the originand diversification of vascular plants through a comprehensive Bayesian analysisof the fossil record. New Phytol. 207: 425-436. Stein, W.E.; Berry, C.M.; Hernick, L.V.A.; Mannolini, F. 2012: Surprisingly complex community discovered in the mid-Devonian fossil forest at Gilboa. Nature 483: 78-81. Stewart, W.N.; Rothwell, G.W. 1993: Paleobotany and the Evolution of Plants. Cambridge University Press: 1-521. Strogen, D.P.; Seebeck, H.; Nicol, A.; King, P.R. 2017: Two-phase CretaceousñPaleocene rifting in the Taranaki Basin region, New Zealand; implications for Gondwana breakup. Journal of the Geological Society, London 174: 929-946. Veen, P.M.V. 1981: Aspects of late Devonian and early Carboniferous palynologyof Southern Ireland. V. The change in composition of palynological assemblages atthe Devonian-Carboniferous boundary. Rev. Palaeobot. Palynol. 34: 67-97. Wang, Y., Xu, H., Fu, Q.; Tang, P. 2004: A new diminutive plant from the Hujiersite Formation (late Middle Devonian) of North Xinjiang, China. Acta Palaeontologica Sinica 43: 461-471. Wang, Y.; Berry, C.M. 2003: A reconsideration of Dimeripteris cornuta Schweitzer and Cai, a diminutive fossil plant from the Middle Devonian of Yunnan, China. Geobios 36: 437-446. Wang, Y.; Berry, C.M.; Hao, S.; Xu, H.; Fu, Q. 2007: The Xichong flora of Yunnan, China: diversity in late Mid Devonian plant assemblages. Geological Journal 42: 339-350. Ward, P.; Labandeira, C.; Laurin, M.; Berner, R.A. 2006: Confirmation of Romer’s Gap as a low oxygen interval constraining the timing of initial arthropod and vertebrate terrestrialization. Proceedings of the National Academy of Sciences of the USA 103 (45): 16818-16822. Xu, H.; Wang, Y.; Tang, P.; Wang, Y. 2017: A new diminutive euphyllophyte from the Middle Devonian of West Junggar, Xinjiang, China and its evolutionary implications. Alcheringa 41 (4): 524-531. Zhu, M.; Yu, X. 2002: A primitive fish close to the common ancestor of tetrapods and lungfish. Nature 418: 767-770. |
| Peripatus Home Page |
Hits counted from 27 Jan 2017:
My Traffic Estimate