| Peripatus Home Page | Updated: 17-Jul-2024 |
AbstractThis page describes the Jurassic Period, including stratigraphy, paleogeography, and famous lagerstðtten, followed by a sketched outline of some of the major evolutionary events.Keywords: Jurassic, Jurassic biota, fossil record, evolution Introduction |
|
||||||||||
StratigraphyHistorical DevelopmentLower (Triassic–Jurassic) Boundary“The end-Triassic mass extinction terminated many groups of marine life, including the conodonts, whose distinctive phosphatic jaw elements constitute a primary zonation for much of the Paleozoic and Triassic, and the majority of ammonoids. Indeed, in the few regions with continuous deposition there is an interval devoid of either typical latest-Triassic taxa (e.g., conodonts or Choristoceras ammonoids) or earliest-Jurassic forms (e.g., Psiloceras ammonites). A sea-level fall produced extended gaps in many shallow-marine sections; therefore, the boundary between upper Triassic and the overlying lower Jurassic was commonly a sequence boundary and hiatus” (Gradstein et al. 2012, p. 733, and references therein). The GSSP for the base of the Jurassic is set at 5.80 m above the base of the Tiefengraben Member of the Kendelbach Formation, corresponding to the local lowest occurrence of the ammonite Psiloceras spelae subsp. tirolicum, in the Kuhjoch section, Northern Calcareous Alps, Austria. Other useful markers include the FAD of Cerebropollenites thiergartii (a pollen grain), Praegubkinella turgescens (a foraminifer), and Cytherelloidea buisensis (an ostracod) (see Gradstein et al. 2012). Upper (Jurassic–Cretaceous) BoundaryThe base of the Cretaceous System is conventionally aligned with the base of the Berriasian Stage; that is, the Tithonian/Berriasian boundary is the Jurassic/Cretaceous boundary. As yet there is no ratified GSSP for the base of the Berriasian Stage although, by common usage, it lies near the first appearance datum (FAD) of the ammonite, Berriasella jacobi. Unfortunately, this ammonite is largely confined to the Mediterranean realm, so the datum is not much use internationally. At various times there have been proposals to redefine the Jurassic/Cretaceous boundary to align with the much more distinct Berriasian/Valanginian boundary (see Énay 2019 for an overview of the argument for such a move). These suggestions did not gain much traction initially, but may be gaining the upper hand, or at least serious consideration, more recently (Granier 2020). ChronologyPaleogeography
|
|||||||||||
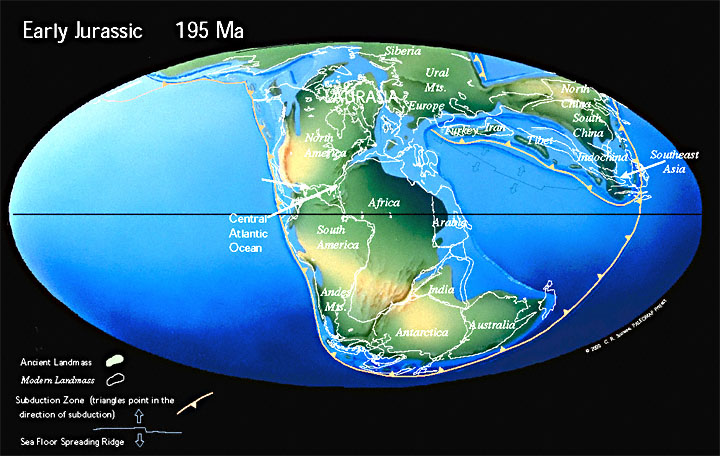
Land and Sea |
(1) |
||||||||||
|
Paleontology
Major TaxaBirdsBird origins have been famously contentious from the earliest days of evolutionary theory. The three most plausible ideas to emerge from the debate are (1) that they evolved from some unknown group of basal archosaurs, probably in the Triassic Period; (2) that they are a sister group to the Crocodylians, perhaps arising from within the sphenosuchian crocodylomorphs in the Early Jurassic; and (3) certainly the most widely held view today, for an ancestry among the theropod dinosaurs, specifically the Maniraptora, in the Middle to early Late Jurassic. Some authors present the origin of feathers as essentially the same phenomenon, on the reasonable premise that feathers are such a unique and diagnostic feature of birds, that it is inconceivable any other group of animals could possess them. In essence, anything with feathers is a bird, anything without them, is not. This view is not unreasonable, but it is not unquestionable either. Despite intensive searching, the earliest known bird is still the famous Archaeopteryx, known from only seven skeletons and an isolated feather, all recovered from the Late Jurassic (Tithonian, ~152 to 145 Ma) Solnhofen Limestone of Germany. The small theropod dinosaur, Compsognathus, has also been recovered from the Solnhofen. This fossil record represents a difficult problem for advocates of the theropod hypothesis: birds (specifically Archaeopteryx) are supposed to be most closely related to the dromaeosaurids, which do not appear in the fossil record until Albian times (mid Cretaceous, about 110 Ma; possibly not – see Wang & Zhou 2017 and update this bit) yet Compsognathus, which is believed to have diverged from the theropod lineage long before the evolution of the dromaeosaurids, occurs alongside Archaeopteryx 40 million years earlier. At present, only the vagaries of the fossil record can be invoked to ‘explain’ the stratigraphic disjunction; our present understanding is unsatisfactory. Mammals“The Jurassic period is an important stage in early mammalian evolution, as it saw the first diversification of this group, leading to the stem lineages of monotremes and modern therian mammals. However, the fossil record of Jurassic mammals is extremely poor, particularly in the southern continents. Jurassic mammals from Gondwanaland are so far only known from Tanzania and Madagascar, and from trackway evidence from Argentina” (Rauhut et al. 2002, p. 165). A study by Meredith et al. (2011) derived the mean date for the split between placentals and marsupials as ~190 Ma, noting that this estimate accords well with the discovery of a stem eutherian from the Jurassic reported by Luo et al. 2011. LagerstðttenPosidonia/Holzmaden Shale: Early Jurassic; Holzmaden, approx. 30 km east of Stuttgart, Germany; fossil reptiles – noteably ichthyosaurs, crustaceans, cephalopods; Selden & Nudds 2012, Hauff & Hauff 1981Stonefield Slates: Middle Jurassic; Stonefield, Oxfordshire, England; bed of sandy slate 30-45 cm thick; surface occurrences exhausted and the underground mine is now closed; most important for a large number of small mammal jaws and teeth representing three orders: Multituberculata, Triconodonta and Pantotheria; also pterosaurs, crocodilians, invertebrates and a possible dicotyledonous angiosperm Christian Malford: Middle Jurassic; England; soft-body preservation of squids; Allison 1988 Solnhofen Limestone: Late Jurassic (Lower Titonian); Altmuhl Valley, Bavaria, Germany; fine-grained lagoonal sediments; most famous for the Archeopteryx and Compsognathus fossils found there, though these two together amount to only nine out the many thousands of specimens known from the Solnhofen; Barthel et al. 1990, Viohl 1985. Morrison Formation: Late Jurassic; North American states of Wyoming and Colorado Purbeck Beds: Late Jurassic; England, esp. near Drulston Bay; essentially modern, temperate insect fauna including dragonflies, locusts, grasshoppers, butterflies, ants and aphids; elsewhere (”Beckel’s Mammal Pit”) numerous mammal species representing five orders and having similarities with those of the contemporaneous Morrison Formation of Wyoming and Colorado
New Zealand OccurrencesFor most of the Paleozoic and Mesozoic, the rocks which would become the basement rocks of the Zealandia continent formed part of the Pacific margin of Gondwana, flanking Australia and Antarctica (Edbrooke 2017; Strogen et al. 2017). During this time, the Zealandia basement developed mainly by subduction-driven, episodic accretion. ReferencesAllison, P.A. 1988: Phosphatised soft bodied squid from the Jurassic Oxford Clay. Lethaia 21: 403-410. Barthel, W.; Morris, S.C.; Swinburne, N.C. 1990: Solnhofen, 2nd ed. Cambridge University Press. Edbrooke, S.W. 2017: The geological map of New Zealand. GNS Science Geological Map 2: 1-183. Énay, R. 2019: The Jurassic/Cretaceous system boundary is an impasse. Why do not go back to Oppel’s 1865 original an historic definition of the Tithonian? Cretaceous Research 106: 104241. Gradstein, F.M.; Ogg, J.G.; Schmitz, M.D.; Ogg, G.M. 2012: The Geologic Time Scale 2012. Elsevier 1-2. Granier, B.R.C. 2020: Introduction to thematic issue, “The transition of the Jurassic to the Cretaceous: an early XXIth century holistic approach”. Cretaceous Research 114: 104530. Hauff, B.; Hauff, R.B. 1981: Das Holzmadenbuch. Self published: 1-136. Luo, Z.; Yuan, C.; Meng, Q.; Ji, Q. 2011: A Jurassic eutherian mammal and divergence of marsupials and placentals. Nature 476: 442-445. Meredith, R.W.; Janečka, J.E.; Gatesy, J.; Ryder, O.A.; Fisher, C.A.; Teeling, E.C.; Goodbla, A.; Eizirik, E.; Simão, T.L.L.; Stadler, T.; Rabosky, D.L.; Honeycutt, R.L.; Flynn, J.J.; Ingram, C.M.; Steiner, C.; Williams, T.L.; Robinson, T.J.; Burk-Herrick, A.; Westerman, M.; Ayoub, N.A.; Springer, M.S.; Murphy, W.J. 2011: Impacts of the Cretaceous terrestrial revolution and KPg extinction on mammal diversification. Science 334: 521-524. Ogg, J.G.; Ogg, G.; Gradstein, F.M. 2008: The concise geologic time scale. Cambridge University Press: 1-177. Rauhut, O.W.M.; Martin, T.; Ortiz-Jaureguizar, E.; Puerta, P. 2002: A Jurassic mammal from South America. Nature 416: 165-168. Nature. Selden, P.; Nudds, J. 2012: Evolution of Fossil Ecosystems (2nd Edition). Academic Press: 1-288. Strogen, D.P.; Seebeck, H.; Nicol, A.; King, P.R. 2017: Two-phase CretaceousPaleocene rifting in the Taranaki Basin region, New Zealand; implications for Gondwana breakup. Journal of the Geological Society, London 174: 929-946. Viohl, G. 1985: Geology of the Solnhofen Lithographic Limestone and the habitat of Archeopteryx. In Hecht, M.K.; Ostrom, J.H.; Viohl, G.; Wellnhofer, P. (ed.) 1985: The beginnings of birds. Eichstatt. Freude des Jura-Museums Eichstðtt, Willibaldsburg: 31-44. Wang, M.; Zhou, Z. 2017: Chapter 1. The evolution of birds with implications from new fossil evidences. In Maina, J.N. 2017: The biology of the avian respiratory system: Evolution, development, structure and function. Springer: 1-26. |
| Peripatus Home Page |
Hits counted from 23 Dec 2017:
My Traffic Estimate