| Peripatus Home Page | Updated: 17-Jul-2024 |
AbstractThis page describes the Ordovician Period, including stratigraphy, paleogeography, and famous lagerstätten, followed by a sketched outline of some of the major evolutionary events.Keywords: Ordovician, Ordovician biota, fossil record, evolution IntroductionThe Ordovician Period is the second period of the Paleozoic Era. This period saw the origin and rapid evolution of many new types of invertebrate animals which replaced their Cambrian predecessors. About 480 million years ago, in the Ordovician period, life forms diversified dramatically and gave rise to many of the marine forms familiar today. The fossil record of this period is amazingly intact in the Great Basin of California, Utah and Nevada and affords an almost unprecedented opportunity to learn about the conditions that favor innovation in biodiversity. Primitive vascular plants appeared the land, until then almost totally barren. The supercontinent of Gondwana drifted over the south pole, initiating a great Ice Age that gripped the earth at this time. The end of the period is marked by a major extinction event. |
|
||||||||||
StratigraphyType Section/SectionsThe type section for the Ordovician, like that of the Cambrian and Silurian, is also in western Wales. The system was founded by Charles Lapworth in 1879, in part as a compromise to resolve the Murchison-Sedgwick conflict over their overlapping claims for their Silurian and Cambrian systems: In 1831 Sedgwick and Murchison commenced work on the stratigraphy of northern Wales. Sedgwick began at the bottom of the section and Murchison at the top. Sedgwick named his sequence of rocks the Cambrian and Murchison applied the name Silurian to the generally more fossiliferous upper formations. Eventually their sections overlapped, each claiming some of the same rocks for their systems, in what became a widespread and lengthy controversy until, forty four years later, Lapworth introduced the name Ordovician for the disputed sequence. Despite this somewhat contrived basis, the Ordovician does comprise a distinct period in Earth history, with a distinctive biota bounded above and below by major extinction events. The name “Ordovician” comes from Ordovices an ancient Celtic tribe that once inhabited the region in Wales where rock strata of this period occur. The Ordovician was originally divided into two sub-periods, Bala and Dyfed, but more recently the Tremadoc epoch was removed from the Cambrian and a three-fold division of Ordovician strata instituted. Lower (Cambrian-Ordovician) BoundaryThe Cambrian was characterised by generally high extinction rates; at least two events are usually ranked among the great Phanerozoic “mass extinctions”. The later of these two events occured at or near the end of the period and notably decimated the trilobites. Steadily accumulating data has led to changing opinions as to where the Cambrian-Ordovician boundary should be formalised. “Traditional concepts of the Cambrian-Ordovician boundary [were] based on the occurrence of the graptolite Dictyonema (formerly Rhabdinopora) flabelliforme (Eichwald 1840), which has a limited regional distribution. Strata with Dictyonema flabelliforme are often difficult to correlate precisely so that different species (and subspecies) of conodonts were favoured for definition of the Cambrian-Ordovician boundary. The Cambrian-Ordovician Boundary Working Group finally decided in 1998 by majority that the base of the Ordovician should be placed at the base of the zone with Iapetognathus fluctivagus, which approximates the Cordylodus lindstromi Zone, the Dictyonema flabelliforme Zone and the FAD of the trilobite Jujuyaspis” (this text formerly available on the International Subcommission on Cambrian Stratigraphy web page). The GSSP for this boundary was chosen at the Green Point section, Newfoundland, approved by the International Commission on Stratigraphy in December 1999 and ratified by the IUGS in January 2000. By a unanimous vote taken in February 2000, the Subcommission selected the name Tremadocian for the lowest stage of the Ordovician System. Upper (Ordovician-Silurian) BoundaryNear Moffat in South Scotland, an uninterrupted Ordovician-Silurian boundary sequence of deep water black shales is exposed at Dobb’s Linn. Here Lapworth (1878) determined the stratigraphy based largely upon graptolites, the dominant macrofossils found in the exposure. Approximately 100 years later, in 1985, this locality was chosen as the International Stratotype for this transition. The boundary is defined at the first appearance of the graptolite Akidograptus ascensus, at the base of the acuminatus Zone, 1.6m above the base of the Birkhill Shales. Follow the scree slope towards the top right hand margin, look for an outcrop which comprises a sequence of five pale-coloured clay bands set in grey mudstone (Upper Hartfell Shale). This is called the anceps Zone. Above this the first appearance of black shales represents the bottom of the Birkhill Shales and the G. persulptus Zone. The Ordovician-Silurian boundary occurs 1.6m above this lithological boundary. ChronologyToday, the International commission on Stratigraphy places the base of the Ordovician at 485.4 ± 1.9 Ma and the top at 443.8 ± 1.5 Ma (Cohen et al. 2015). PaleogeographyMajor Tectonic EventsIn the latest Precambrian and early Paleozoic, the supercontinent Rodinia, centered about the south pole, broke apart as blocks drifted northward. Most notable of these blocks were the large continents North America [Laurentia], Baltica, and Siberia.During the Ordovician ancient oceans separated the barren continents of Laurentia, Baltica, Siberia and Gondwana. The end of the Ordovician was one of the coldest times in Earth history. Ice covered much of the southern region of Gondwana. Taconian Orogeny (~475 Ma) – Numerous plates and continental blocks approach North America from the south and east. |
|||||||||||
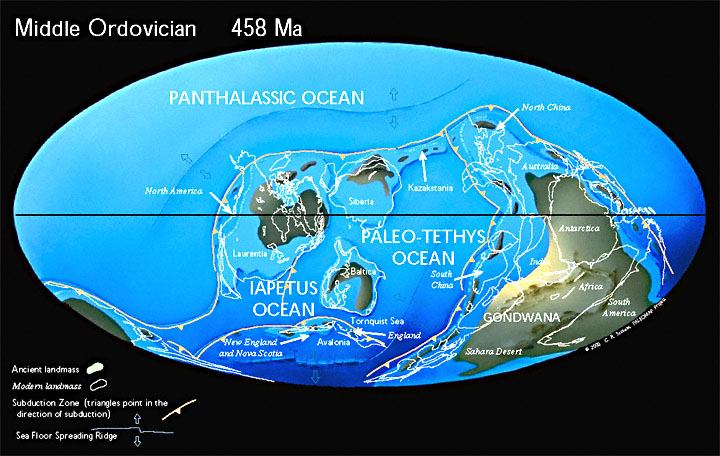
Land and SeaDuring the Ordovician, Southern Europe, Africa, South America, Antarctica and Australia remained joined together into the supercontinent of Gondwanaland, which had moved down to the South Pole. North America straddled the equator, and was about 45 degrees clockwise from its present orientation. Western and Central Europe were separate from the rest of Eurasia, and were rotated about 90 degrees counterclockwise from their present orientation, and was in the southern tropics. North America is engaged in a slow collision with the microcontinent of Baltica, which forms the core of what is later to become Europe. The Iapetus Ocean continues to shrink as the previously passive margins of Baltica and North America converge. Where the Iapetus was, mountains are thrust up, remnant strata of which remain today in Greenland, Norway, Scotland, Ireland and north-eastern North America. Scotland and England are united into a single landmass.Major transgression in middle Ordovician, therefore widespread shallow, warm epeiric seas. |
(1) |
||||||||||
ClimateThe early Ordovician is thought to have been relatively warm; extensive coral reefs were developed in the tropics. Elevated carbon dioxide levels through the Cambrian and early Ordovician may have greenhouse conditions with global summer temperatures as high as 40°C (<$catg ref Crowley & North 1991>). “However, by the late Ordovician (~458-443 Ma) there is evidence to suggest that global climates had become much more variable, and that certain regions were becoming cool and moist” (<$catg ref Willis & McIlwain 2002>). There is evidence for a rapid drop in sea level in the Late Ordovician and climatic cooling leading to a major glaciation in Gondwana near end of the period.“Paleoclimate records indicate global cooling during ~35 m.y. of the Early and Middle Ordovician, a relatively stable climate in the ~8-m.y.-long Katian Age, and a glacial maximum in the Hirnantian Age (445.2–443.8 Ma).... Cooling likely resulted in the establishment of ice sheets on polar Gondwana in the Katian Age or even earlier.... The subsequent rapid expansion of Hirnantian ice sheets resulted in glacioeustatic sea-level fall and decreased temperatures … associated with the first pulse of the Late Ordovician mass extinction (LOME).... The second pulse of the LOME occurred as glaciers melted, oceans warmed, and anoxic seawater transgressed the shelves.... The cause of the glacial advance is unknown, but has been attributed to CO2 levels falling below a critical threshold...” (Jones et al. 2017, p. 631). As Gondwana moved south, Africa, now straddling the south pole, became extensively glaciated. There were even glaciers in what is now the Sahara. The glaciation caused global temperatures to drop as the period progressed, and the world entered an ice age, although conditions remain mild and equitable in the tropics. “In terms of abiotic changes, a series of secular, deterministic changes occurred to modify the marine environment from a late Cambrian world characterized by episodic anoxia, extremely high ocean temperatures, and sluggish ocean circulation to a mid-Ordovician world characterized by lower temperatures, cyclical sea level changes, oceanic ventilation and higher levels of atmospheric oxygen, and fully circulating oceans” (Stigall et al. 2019, p. 266). PaleontologyIntroductionThe Ordovician was an age of evolutionary experimentation, in which new organisms evolve to replace those that died out at the end of the Cambrian. It was one of the largest adaptive radiations in the Earth’s history. The number of families of known marine invertebrates increases from about 200 at the end of the Cambrian to around 500 in the early Ordivician. The widespread shallow, warm continental seas were the perfect environment for many groups of organisms. Microorganisms such as colonial blue-green algae – stromatolites – are widespread. Foraminifera evolve. Acritarchs, although existing during the Precambrian, become more common. Stromatoporoids (possibly sponge-like organisms) also appear.Essentially ‘modern’ (in the sense that problematica are mostly absent and all extant high level taxa represented) marine invertebrate fauna; however, land remains barren and vertebrates undeveloped. “Diversity of marine life reached a peak during the Ordovician period. Most newly arising species represent modifications of existing forms, many of which made their first appearance during the preceding Cambrian period. Whereas many of the dominant Cambrian life forms waned or became extinct, some of the new groups that appeared during the Ordovician dominated the next 250 million years of the Paleozoic era. For example, the now-extinct graptolites first appeared in the Cambrian, but diversified dramatically during the Ordovician. In addition, many Ordovician brachiopods, mollusks, corals and stelleroids bear some resemblance to their modern descendants” (Droser et al. 1996). An interesting phenomenon is the sudden increase in filter feeding organisms. Cambrian animals were predominately crawling mud-grubbers and detrivores with a few swimming and burrowing predators thrown in. Filter feeders (such as Dinomischus and Lepidocystis) although an important part of the fauna, were not exceedingly common. In the Ordovician an increase in the amount of micro-plankton would be the obvious explanation for the sudden increase in number and diversity of filter-feeders. Groups absent or under-represented in the Cambrian suddenly become more important. We see the first appearance of the corals, including both rugose (solitary) and tabulate (colonial) forms, bivalve molluscs, and the planktonic graptolites (Graptoloidea). The lophophorates (animals that suck food from the water using a special fringe of tentacles around the mouth) are more filter feeders that do well in the Ordovician. The Bryozoa appear in large numbers, and constitute the most predominant colonial animals of the time. Each bryozoan polyp is a tiny creature, not unlike a coral animal. On the continental shelves of the isolated and drifting continents, marine organisms engage in evolutionary experiments. There are two main geographical provinces, a northern, equatorial tropical one, and a southern cool-water one centered around Gondwanaland. In the case of trilobites, North America and northwestern margins of Europe, Spitzbergen, Siberia and north-east Russia are characterised by a diverse fauna that H. B. Whittington calls the Bathyurid fauna. This includes trilobites of the families Bathyuridae, Hystricuridae, Asaphidae, Komaspididae, Remopleurididae and Pliomeridae. A subprovince around Baltica bathyurids and pliomerids are rare and asaphids belong to different genera, indicated the Iapetus Ocean remained wide enough to act as a barrier to migration for these shallow water continental-shelf forms. In the cooler waters of Gondwanaland are found the Selenopeltis and Hungaiid-Calymenid faunal provinces, including trilobites of the families Hungaiidae, Calymeniidea, Pliomeridae, Illaenidae, and endemic Asaphidae. The differences between Bathyurid fauna and Hungaiid-Calymenid faunal provinces were due to a combination of climate factors and geographical separation. The same biogeographical distribution seems to apply to articulate brachiopods as well. The Balto-Scandian region is better defined in the brachiopod then the trilobite faunas, and for much of the period can be considered a distinct biogeographical province. In the later part of the Ordovician genera of families previously limited to one faunal province appear in another, indicating a tendency towards migration and cosmopolitism. Changed in oceanic circulation (distribution of planktonic larvae etc) along with approaching continental masses would have made possible migrations of shallow water benthos. Major Evolutionary EventsThe Ordovician was a time of adaptive radiation of many faunal groups, following the mass extinction of trilobites and nautiloids at end of Cambrian. “Since the earliest comprehensive compilations of Phanerozoic marine biodiversity, a rapid and pronounced rise in taxonomic richness has been recognized during Ordovician Period (Valentine 1969).... Notably, with the advent of Jack Sepkoski’s kinetic model of taxonomic diversity, a particularly clear trend emerged of a massive taxonomic radiation at various levels that led to significant accumulation of biodiversity centered on the Middle Ordovician. … The terms Ordovician Radiation (Sepkoski & Sheehan 1983) and Great Ordovician Biodiversification Event (Webby et al. 2004) [→ sidebar] both refer to the dramatic radiation of marine biodiversity that occurred between the late Cambrian Furongian crisis interval and Late Ordovician Mass Extinction. During this interval, present ordinal-level diversity of metazoans was achieved, family-level diversity increased threefold, and genus level diversity more than quadrupled. Furthermore, the profound faunal transition from the Cambrian to Paleozoic Evolutionary Faunas (EF) occurred, primarily through the expansion of the level-bottom suspension feeding benthos” (Stigall et al. 2019, p. 249-250).The base of the North American Whiterock Series coincides with the start of the great Ordovician radiations of articulate brachiopods, bryozoans, bivalves, echinoderms, and virgellinid graptoloids. Many other groups first appear in the fossil record at or near this horizon. This also marks the point of transition between dominance of Sepkoski’s Cambrian Evolutionary Fauna, of which trilobites are the main component, and the rise of the Paleozoic Evolutionary Fauna composed of the radiating groups. However, the Whiterock trilobite fauna shows explosive and sustained radiation at exactly this point, conforming to the Paleozoic fauna diversity pattern, and indicating that trilobites were active participants in the Ordovician radiation. |
|
||||||||||
Major TaxaRepresentative fauna:brachiopods, rugose & tabulate corals, bryozoans, crinoids, conodont animals, jawless fishes Most marine organisms were epifaunal; few infaunal Predators Evolution of increasing effective predators. Major predators were invertebrates Nautiloids Starfish Decline of stromatolites New kinds of reefs appeared bryozoans, stromatoporoids, tabulate corals The armoured placoderms began to differentiate from the main gnathostome lineage in the Ordovician, followed by the chondrichthyans (sharks and rays), also in the Ordovician. Plants probably colonized the land; land plants were well established by the Silurian. Corals (subclass Zoantharia) were represented in the Cambrian by the Cothoniida and some tabulate-like corals, though the earliest undoubted Rugosa and Tabulata are Ordovician. By the Middle Ordovician the full range of colonial tabulate types had arisen, and they remained more numerous than the Rugosa throughout the Ordovician. However, the Tabulata were more profoundly affected by the Late Ordovician extinction. Among the molluscs were newcomers such as bivalves, which were not common during this time, although the late Cambrian Archaeogastropods develop at a moderate pace. Virtually all the major bivalve stocks were established by the Middle Ordovician (Clarkson 1993, p. 205). A much more spectacular success story were the nautiloid cephalopods. Small and rare in the late Cambrain, the nautiloids evolve quickly along many different lines. At least ten different orders flourished at this time, all but one appearing for the first time during the early or middle part of the Ordovician. This astonishing diversity included straight, curved, loosley coiled, and tightly coiled shelled types, and even one group (the Ascocerids) that in order to become lighter and more streamlined lost the a large part of their shell altogether. These carnivorous molluscs replaced the Cambrian Anomalocarids as the dominant life form and top predator of the world’s ocean. The biggest, such as the endocerids, attained huge size; with shells 3 to 5 meters or more in length they were the largest animal that, up until that time, had ever lived. Ordovician trilobites were for the most part quite different from their Cambrian predecessors. Many evolved bizarre spines and nodules. Others, such as Aeglina prisca from the warm shallow seas of what is now Bohemia, were clearly pelagic nektonic swimming forms, and developed huge eyes with over a thousand facets, while still others went the other direction and lost their eyes altogether. Some trilobites developed shovel-like snouts for ploughing through mud, others fused the segments of their bodies. The curious Trinucleids developed a broad pitted margin around the head shield. The ‘bryozoans’ are thought (e.g. Nielsen 2001, p. 232-234) to be a paraphyletic group, so the entoprocts and ectoprocts should be treated separately. In any event, it is only the ectoprocts which appear in the fossil record as early as the Ordovician; one of the few modern phyla which did not become established until after the Cambrian. After humble Cambrian beginnings the articulate brachiopods greatly increase in diversity and abundance, with no fewer than fourteen new superfamilies. The Orthid and Strophomenid orders were especially diverse. A number of early echinoderm experiments die out, others straggle on, while still others increase in diversity. The echinoderms included both a great many stalked (filter-feeding) and a few mobile (some predatory) forms. Among the bizarre forms were the carpoids, which were able to push themselves along the mud by means of a stout “tail.” During the Ordovician the Crinoids, rare during the Cambrian, suddenly appear and diversify in large numbers. Like the brachiopods these sessile benthic (attached bottom-living) invertebrates were to become an important group of filter feeders throughout the rest of the Paleozoic. The echinoderm classes living today, except the holothuroids, are known from the early Ordovician, while all other classes of the Cambrian radiation became extinct during the Paleozoic. (After Nielsen 2001, pp. 419-420.) The Ordovician was the high point of the graptolites, particularly the planktonic Graptoloidea. These evolved from benthic attached late Cambrian forms and diversified in earliest Ordovician (Tremadoc) time into a number of different planktonic types, including single-branched single and double rowed colonies, two-branched, four-branched, and spiral forms. In New Zealand, Ordovician anoxic deep sea black shales with abundant graptolites are well represented in North West Nelson (Cooper 1979). The vertebrate ostracoderms remain rare, although several different groups evolve. Their cousins the conodont animals, worm-like or eel-like organisms known mostly from numerous isolated denticles (which were used to support some kind of grasping or breathing structure in the mouth or throat) represent a major component, quite possibly predators and certainly nektonic/pelagic, in the marine food-chain. “The oldest identifiable gnathostome [jawed fishes] fossils are extremely scrappy, consisting of isolated scales and teeth. Because paleontologists find similar scales and teeth in more recent fossils that are complete enough to reveal the presence of jaws, it is inferred that the early fishes from which these bony fragments came also had jaws. Among the most ancient scraps are supposed shark scales from the Silurian of Mongolia and the Ordovician of the USA, thought to be about 420 and 450 million years old respectively, resembling the simple skin denticles (small toothlike scales) of modern sharks” (Maisey 1996, p. 61). During the Ordovician the first creeping lichens and bryophytes move onto land, the beginning of a great new experiment of life.
ExtinctionsMass extinctions of tropical marine faunas occurred at the end of the Ordovician when 100 or more families became extinct, including more than half of the bryozoan and brachiopod species.Possible causes which have been mooted include:
New Zealand OccurrencesFor most of the Paleozoic and Mesozoic, the rocks which would become the basement rocks of the Zealandia continent formed part of the Pacific margin of Gondwana, flanking Australia and Antarctica (Edbrooke 2017; Strogen et al. 2017). During this time, the Zealandia basement developed mainly by subduction-driven, episodic accretion. The Ordovician is not widely represented in New Zealand, although there are a few occurences, mainly in the north west Nelson area (see Cooper 2004). ReferencesClarkson, E.N.K. 1993: Invertebrate Paleontology and Evolution (third edition). Chapman and Hall. Cohen, K.M.; Finney, S.C.; Gibbard, P.L.; Fan, J.X. 2015: The ICS international chronostratigraphic chart v 2015/01. Episodes 36: 199-204. Cooper, R.A. 1979: Ordovician Geology and Graptolite Faunas of the Aorangi Mine Area, North West Nelson, New Zealand. New Zealand Geological Survey Paleontological Bulletin 47: 1-127. Cooper, R.A. (ed.) 2004: The New Zealand geologic timescale. Institute of Geological and Nuclear Sciences Monograph 22: 1-284. Edbrooke, S.W. 2017: The geological map of New Zealand. GNS Science Geological Map 2: 1-183. Eichwald, C.E. 1840: Über das silurische Schichtensystems in Esthland. Zeitschrift für Natur-und Heilkunde der K. Medicinisch-chirurgische Akademie St. Petersburg 1/2: 1-210. Jones, D.S.; Martini, A.M.; Fike, D.A.; Kaiho, K. 2017: A volcanic trigger for the Late Ordovician mass extinction? Mercury data from south China and Laurentia. Geology 45 (7): 631-634. Lapworth, C. 1878: The Moffat Series. Quarterly Journal of the Geological Society of London 34: 240-346. Nielsen, C. 2001: Animal evolution: Interrelationships of the living phyla (second edition). Oxford University Press: 1-378. Ogg, J.G.; Ogg, G.; Gradstein, F.M. 2008: The concise geologic time scale. Cambridge University Press: 1-177. Sepkoski Jr., J.J. 1995: The Ordovician Radiations: diversification and extinction shownby global genus level taxonomic data. In Cooper, J.D.; Droser, M.L.; Finney, S.C. (ed.) 1995: Ordovician Odyssey: Short Papers. 7th International Symposium on the Ordovician System. Society for Sedimentary Geology (SEPM) 77: 393-396. Sepkoski Jr., J.J.; Sheehan, P.M. 1983: Diversification, faunal change, and community replacement during the Ordovician Radiations. In Tevesz, M.J.S.; McCall, P.L. (ed.) 1983: Biotic Interactions in Recent and Fossil Benthic Communities. Plenum Press, New York: 673-717. Stigall, A.L.; Edwards, C.T.; Freeman, R.L.; Rasmussen, C.M.Ø. 2019: Coordinated biotic and abiotic change during the Great Ordovician Biodiversification Event: Darriwilian assembly of early Paleozoic building blocks. Palaeogeography, Palaeoclimatology, Palaeoecology 530: 249-270. Strogen, D.P.; Seebeck, H.; Nicol, A.; King, P.R. 2017: Two-phase Cretaceous–Paleocene rifting in the Taranaki Basin region, New Zealand; implications for Gondwana breakup. Journal of the Geological Society, London 174: 929-946. Valentine, J.M. 1969: Patterns of taxonomic and ecological structure of the shelf benthos during Phanerozoic time. Palaeontology 12: 684-709. Webby, B.D., Paris, F., Droser, M.L., Percival, I.G. (ed.) 2004: The Great Ordovician Biodiversification Event. Columbia University Press. |
| Peripatus Home Page |
Hits counted from 10 May 2017:
My Traffic Estimate