| Peripatus Home Page | Updated: 12-Jul-2025 |
AbstractThis page describes the Permian Period, including stratigraphy, paleogeography, and famous lagerstätten, followed by a sketched outline of some of the major evolutionary events.Keywords: stratigraphy, Permian Period, Permian biota, fossil record, evolution, extinction, Wellington Shale IntroductionThe Permian Period began with drastic climatic changes resulting from the convergence of Laurentia, Eurasia and Gondwana to form the supercontinent of Pangea, and ended with the most profound mass extinction event known. The lush clubmoss and horsetail forests which formed the vast Carboniferous coal measures were mostly replaced by plants adapted to drier conditions and, in the southern hemisphere, by the iconic Glossopteris seed-fern forests. Tetrapods diversified, notably giving rise to the therapsids from which later arose the mammals. |
|
||||||||||
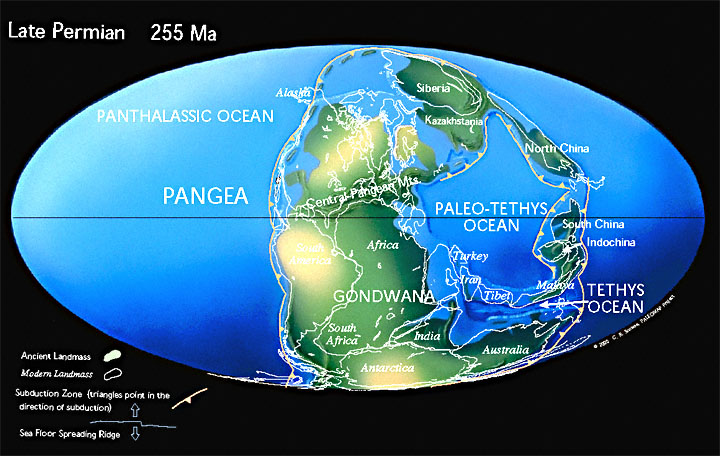
StratigraphyHistorical DevelopmentThe Permian was established by Murchison (1841) and named after the “ancient kingdom of Permia that formerly lay between the Volga River and the Ural mountains in Russia” (Ogg et al. 2008, p. 85). Lower (Carboniferous–Permian) BoundaryThe GSSP defining the the base of the Permian (i.e., the Carboniferous-Permian Boundary) was established in 1998 (Davydov et al. 1998) at Aidaralash Creek, Kazakhstan, at the first appearance of the “isolated-nodular” morphotype of the conodont Streptognathodus “wabaunsensis”. Upper (Permian–Triassic) BoundaryThe mass extinction event which marks the Permian-Triassic boundary is discussed elsewhere. Stratigraphically, the boundary is defined at the first occurrence of the conodont Hindeodus (al. Isarcicella) parvus within the evolutionary lineage Hindeodus typicalise – H. Latidentatus praeparvuse – H. parvuse – H. postparvus as the primary correlation marker for the base of the Mesozoic and Triassic (Gradstein et al. 2012, p. 683). ChronologyToday, the Permian is considered to range from 298.9 ± 0.15 Ma to 252.17 ± 0.06 Ma (Cohen et al. 2015). PaleogeographyMajor Tectonic EventsThe ocean separating Siberia and Baltica closed and these two continents collided, completing the assembly of Pangea, and with the ensuing orogeny creating the Ural Mountains. At about the same time or immediately after, Gondwana was shedding peripheral “microcontinents” which would eventually form Tibet and Malaysia. |
|||||||||||
Land and SeaGondwana was probably located over or near the South Pole at the beginning of Permian time, as evidenced by fossil plant groups typical of cool climates. Much of Laurentia was covered by shallow seas, resulting in the deposition of the reef limestones found in present day western Texas. Eurasia, Laurentia and Gondwana merged to form the supercontinent of Pangea, with prfound effects upon the global climate. Northward movement of the former Gondwana landmass diminished the late Carboniferous icecap over the southern part of that region, and the continental interior at low latitudes became hotter and drier. |
(1) |
||||||||||
ClimateThe extensive southern ice cap which formed during the late Carboniferous and persisted through into early Permian times was probably melted during the mid Permian as global temperatures rose.“Overall, the final 5 m.y. of the Permian period in the high southern latitude Bowen Basin [Queensland, Australia] records the end of the final P4 Glaciation of the late Paleozoic Ice Age, followed by continued cold (but nonglacial) conditions, until a final warming trend that began about 253 Ma and continued across the” end-Permian extinction (Fielding et al. 2022, p. 26). PaleontologyIntroductionCarboniferous coal forests diminished in response to increasing aridity. Conifers radiated and spread, particularly in the northern hemisphere; Glossopteris forests dominated in the south.
Major Taxabrachiopodsamphibians mammal-like reptiles LagerstättenBears Ears National Monument: Utah; late Paleozoic (Late Carboniferous to Permian; approximately 315-280 Ma) vertebrate fossil locality; taxa present include freshwater sharks, other fish, amphibians and reptiles, the most common being Eryops and the mammal-like reptile, Ophiacodon. An adjacent Triassic age vertebrate bed, particularly known for phytosaurs, has recently been removed from the National Monument and lost its protected status.Wellington Shale: Near Elmo, Kansas; contains insects (Early Permian) Karoo System: Southern Africa; various taxa, most famously a rich mammal-like reptile fauna from the Beaufort Sandstone (~middle Permian to near the end of the Triassic). ExtinctionsThe Permian was terminated by the greatest known of all extinction events: Of all Phanerozoic mass extinctions, the event at the end of the Permian was by far the most profound. At this time something like 60% of all species (and perhaps 90% of all marine forms) went extinct. The last few trilobites disappeared during this event although, as a class, they were already a very impoverished group by then.No clear explanation is available and it is possible that the extinction was an emergent consequence of many factors which, individually, may have been too subtle to leave much evidence. |
|||||||||||
|
“Fundamental to understanding its cause is determining the tempo and duration of the extinction. Uranium/lead zircon data from Late Permian and Early Triassic rocks from south China place the Permian-Triassic boundary at 251.4 ± 0.3 million years ago. Biostratigraphic controls from strata intercalated with ash beds below the boundary indicate that the Changhsingian pulse of the end-Permian extinction, corresponding to the disappearance of about 85 percent of marine species, lasted less than 1 million years. At Meishan, a negative excursion in d13C [see Fig. 2] at the boundary had a duration of 165,000 years or less, suggesting a catastrophic addition of light carbon” (Bowring et al. 1998, p. 1039).
However, widespread “basaltic volcanism occurred in the region of the West Siberian Basin in central Russia during Permo-Triassic times. New 40Ar/39Ar age determinations on plagioclase grains from deep boreholes in the basin reveal that the basalts were erupted 249.4 ± 0.5 million years ago. This is synchronous with the bulk of the Siberian Traps, erupted further east on the Siberian Platform. The age and geochemical data confirm that the West Siberian Basin basalts are part of the Siberian Traps and at least double the confirmed area of the volcanic province as a whole. The larger area of volcanism strengthens the link between the volcanism and the end-Permian mass extinction” (Reichow et al. 2002, p. 1846, Abstract). “The marine sediments of Jameson Land, East Greenland, preserve a unique record of the Permian-Triassic extinction event. High rates of sedimentation, controlled by active faulting, have produced a greatly expanded succession compared to other sections worldwide. In addition, the sediments have suffered remarkably little burial and thermal alteration and contain well preserved marine organisms and terrestrially derived plant remains. For the first time, it is possible to compare the terrestrial and marine fossil records of this extinction event using the same samples from the same sections. These studies have importance for palaeoecology as well as for correlation between marine and terrestrial sections worldwide. Marine ecosystem collapse is signalled by a sharp reduction in bioturbation, a disappearance of Permian taxa, a sharp decrease in the 13C curve (interpreted as an indication of productivity collapse) and the appearance of widespread oxygen restriction. This all occurs within approx. 50 cm at the top of the Schuchert Dal Formation. During the same interval, there is also a turnover in the terrestrial ecosystem from gymnosperm to pteridosperm dominated floras. Studies of sections in northern Italy also suggest that a similar synchronicity exists in the timing of ecosystem recovery in both marine and terrestrial realms during the late Lower Triassic” (Looy & Twitchett 1999). “There is a widespread belief that ‘throughout their history fish have proved virtually immune to mass extinction events’” (Hallam & Wignall 1997, p. 71). During the Permian-Triassic extinction event fish diversity seems to have increased, whereas all other marine groups declined sharply. This presents an ecological problem: how can organisms at the top of a food chain survive a widespread collapse in productivity and biomass at lower trophic levels? Good Permian-Early Triassic fish faunas are known only from a few areas worldwide: Madagascar, East Greenland and Spitsbergen. One possibility is that in these areas the extinction was less severe than elsewhere and thus the food chain remained relatively intact. However, palaeoecological data from East Greenland show that conditions were just as bad here as elsewhere. Instead, it appears that fish were better preserved in the Lower Triassic than in the latest Permian. This is due to widespread benthic anoxia, high sedimentation rates and rapid concretion formation in the narrow, fault controlled basin of East Greenland. In contrast, benthic invertebrate groups show much worse preservation in the Lower Triassic than in the Late Permian (due to a dearth of silicified faunas). When the number of Lazarus taxa are taken into account, fish diversity shows a similar pattern through the Permian-Triassic interval as all other marine groups” (Twitchett 1999). An analysis of “global Permian and Triassic plant data in a paleogeographic context show that the scale and timing of effects [on plants] varied markedly between regions” and that “the patterns are best explained by differences in geography, climate, and fossil preservation, not by catastrophic events” (Rees 2002, p. 827). |
 Fig. 2. Carbon isotope curve at Meishan section; from Bowring et al. 1998. |
||||||||||
New Zealand OccurrencesFor most of the Paleozoic and Mesozoic, the rocks which would become the basement rocks of the Zealandia continent formed part of the Pacific margin of Gondwana, flanking Australia and Antarctica (Edbrooke 2017; Strogen et al. 2017). During this time, the Zealandia basement developed mainly by subduction-driven, episodic accretion. The Permian is restricted to relatively few localities in New Zealand, mostly in the southern South Island, but over the years a diverse fauna (e.g., Waterhouse 1964) and flora (e.g., Crosbie 1985), including rare Glossopteris leaves (Mildenhall 1970, 1976), have been reported. ReferencesBowring, S.A.; Erwin, D.H.; Jin, Y.G.; Martin, M.W.; Davidek, K.; Wang, W. 1998: U/Pb zircon geochronology and tempo of the end-Permian mass extinction. Science 280: 1039-1045. Cohen, K.M.; Finney, S.C.; Gibbard, P.L.; Fan, J.X. 2015: The ICS international chronostratigraphic chart v 2015/01. Episodes 36: 199-204. Crosbie, Y.M. 1985: Permian palynomorphs from the Kuriwao Group, Southland, New Zealand. New Zealand Geological Survey Record 8: 109-119. Davydov, V.I.; Glenister, B.F.; Spinosa, C.; Ritter, S.M.; Chernykh, V.V.; Wardlaw, B.R.; Snyder, W.S. 1998: Proposal of Aidaralash as Global Stratotype Section and Point (GSSP) for base of the Permian System. Episodes 21 (1): 11-18. Edbrooke, S.W. 2017: The geological map of New Zealand. GNS Science Geological Map 2: 1-183. Fielding, C.R.; Frank, T.D.; Savatic, K.; Mays, C.; McLoughlin, S.; Vajda, V.; Nicoll, R.S. 2022: Environmental change in the late Permian of Queensland, NE Australia: The warmup to the end-Permian Extinction. Palaeogeography, Palaeoclimatology, Palaeoecology 594 (110936). Gradstein, F.M.; Ogg, J.G.; Schmitz, M.D.; Ogg, G.M. 2012: The Geologic Time Scale 2012. Elsevier 1-2. Hallam, A.; Wignall, P.B. 1997: Mass extinctions and their aftermath. Oxford University Press. Looy, C.V.; Twitchett, R.J. 1999: Synchronous collapse, and recovery, of the terrestrial and marine ecosystems during the Permian-Triassic interval. Palaeontological Association 43rd Annual Meeting, University of Manchester, 19-22 December 1999 (Oral Presentation). Mildenhall, D.C. 1970: Discovery of a New Zealand member of the Permian Glossopteris flora. Australian Journal of Science 32 (12): 474-475. — 1976: Glossopteris ampla Dana from New Zealand Permian sediments. New Zealand Journal of Geology and Geophysics 19 (1): 130-132. Murchison, R.I. 1841: First sketch of the principal results of a second geological survey of Russia. Philosophical Magazine, series 3, no. 19: 417-422. Ogg, J.G.; Ogg, G.; Gradstein, F.M. 2008: The concise geologic time scale. Cambridge University Press: 1-177. Rees, P.M. 2002: Land-plant diversity and the end-Permian mass extinction. Geology 30 (9): 827-830. Reichow, M.K.; Saunders, A.D.; White, R.V.; Pringle, M.S.; Al'Mukhamedov, A.I.; Medvedev, A.I.; Kirda, N.P. 2002: 40Ar/39Ar Dates from the West Siberian Basin: Siberian Flood Basalt Province doubled. Science 296: 1846-1849. Strogen, D.P.; Seebeck, H.; Nicol, A.; King, P.R. 2017: Two-phase Cretaceous–Paleocene rifting in the Taranaki Basin region, New Zealand; implications for Gondwana breakup. Journal of the Geological Society, London 174: 929-946. Twitchett, R.J. 1999: Palaeoecology, preservation and the myth that fish are immune to mass extinction events. Palaeontological Association 43rd Annual Meeting, University of Manchester, 19-22 December 1999 (Oral Presentation). Waterhouse, J.B. 1964: Permian stratigraphy and faunas of New Zealand. New Zealand Geological Survey Bulletin 72: 1-111. |
| Peripatus Home Page |
Hits counted from 22 Jun 2017:
My Traffic Estimate