| Peripatus Home Page | Updated: 14-Mar-2024 |
AbstractThis page describes the Silurian Period, including stratigraphy, paleogeography, and the fossil record. Four famous lagerstðtten the Much Wenlock Limestone, Fiddlers Green, Waukesha and Lesmahagow formations are briefly discussed, followed by a sketched outline of some of the major evolutionary events.Keywords: stratigraphy, Silurian Period, Silurian biota, fossil record, evolution, extinction, Wenlock Limestone, Fiddler's Green Formation IntroductionThe first coral reefs became widespread during the Silurian, creating new marine habitats. Some are preserved at the iconic fossil sites in the Wenlock Limestone of Shropshire. Eurypterids were at their peak. Fish diversified, evolving lower jaws and invading fresh water environments. On land, lycopsid plants appeared and early terrestrial ecosystems bacame established. |
|
||||||||||
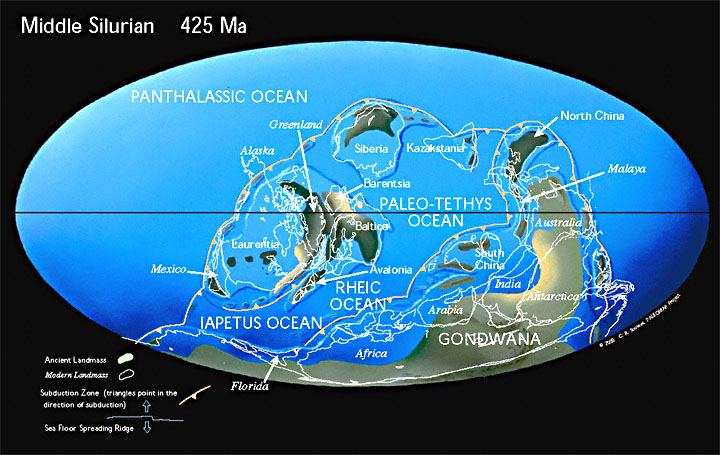
StratigraphyType Section/SectionsIn 1831 Adam Sedgwick and a collaborator, Sir Roderick Impey Murchison, commenced work on the stratigraphy of northern Wales. Sedgwick began at the bottom of the section and Murchison at the top. Sedgwick named his sequence of rocks the Cambrian and Murchison applied the name Silurian to the generally more fossiliferous upper formations, with a type section in western Wales defined both by lithology and fossils (Murchison 1835). Eventually their sections overlapped, each claiming some of the same rocks for their systems, in what became a widespread and lengthy controversy until, many years later, Charles Lapworth (Lapworth 1879) put forward his proposal for the tri-partite division of the lower Paleozoic, introducing the name Ordovician for the disputed sequence (Hamilton 2001). Lower (Ordovician-Silurian) BoundaryNear Moffat in South Scotland, an uninterrupted Ordovician-Silurian boundary sequence of deep water black shales is exposed at Dobb’s Linn. Here Lapworth (1878) determined the stratigraphy based largely upon graptolites, the dominant macrofossils found in the exposure. Approximately 100 years later, in 1985, this locality was chosen as the International Stratotype for this transition. The boundary is defined at the first appearance of the graptolite Akidograptus ascensus, at the base of the acuminatus Zone, 1.6m above the base of the Birkhill Shales. Follow the scree slope towards the top right hand margin, look for an outcrop which comprises a sequence of five pale-coloured clay bands set in grey mudstone (Upper Hartfell Shale). This is called the anceps Zone. Above this the first appearance of black shales represents the bottom of the Birkhill Shales and the G. persulptus Zone. The Ordovician-Silurian boundary occurs 1.6m above this lithological boundary. Upper (Silurian-Devonian) BoundaryThe base of the Devonian is defined immediately at the first appearance of the graptolite species Monograptus uniformis in the rhythmically alterating limestones and calcareous shales of ‘Bed 20’ in the Klonk Section, which is located near the village of Suchomasty, about 35km southwest of Prague in the Czech Republic. The age is established at 419 ± 3.2 Ma. ChronologyToday, the International commission on Stratigraphy places the base of the Silurian at 443.8 ± 1.5 Ma and the top at 419.2 ± 3.2 Ma (Cohen et al. 2015). Subdivisions of the Silurian are primarily provided by the FADs (first appearance data) of graptolite species. PaleogeographyMajor Tectonic Events“During the middle and late Paleozoic, about a third of the Rodinian mass was torn apart and moved to equatorial regions. Most of these blocks were assembled by a series of plate collisions into the super continents of Laurussia [the Old Red continent] by the Devonian and Laurasia by the Pennsylvanian. Meanwhile the remains of Rodinia, Gondwana, rotated clockwise and moved northward to collide with Laurasia -- the result was the super continent Pangaea [all land]. Pangaea was shaped like a huge “pack man” [sic], mouth agape and facing eastward across the equator. The large,open mouth was the Tethys Ocean.Baltica and the attached micro continent Avalonia begin colliding with North America in scissors fashion [north to south] to form the Caledonian-Acadian orogeny” (from a discontinued web page entitled “Global History Silurian” hosted by Northern Arizona University, author unknown). no major volcanic events Taconian Orogeny – Early Silurian; North America Caledonian Orogeny – Late Silurian-Devonian subduction closes northern Iapetus Ocean under Europe; mountain building in northwestern Europe Acadian Orogeny – Late Silurian-Devonian subduction in eastern North America, from Canada to Pennsylvania; ending in collision between Laurentia and Baltica |
|||||||||||
Land and Seadeglaciation => worldwide high sea levels following the Late Ordovician low sea level; generally low continental elevationssea levels dropped again in Late Silurian (?) vast northern polar ocean Rodinia largely broken up; some of the fragments equatorial and eventually reassemble (when?) into Laurussia (sometimes known as Euamerica) and Laurasia Laurentia – Taconic mountains eroding; Appalachians erode down to base by beginning of the Devonian oceans between Laurentia, Baltica and Avalonia continue closing and become substantially closed in the Silurian; collision of Laurentia and Baltica to form Laurussia formation of evaporite deposits in Ludlow/Pridoli times – northern Europe, Siberia, South China, Australia Gondwana over South Pole, ringed by ~6 continents |
(1) |
||||||||||
Climatestable global climate following erratic fluctuations (in Ordovician presumably ?)relatively warm – no large land mass over poles deglaciation; consequent general rise in sea levels beginning of drop in global CO2 levels (which accelerates greatly in the Late Devonian) PaleontologyIntroductionFirst coral reefs widespread; eurypterids at their peak; fish diversify, evolving lower jaws and invading fresh water environments; lycopsid land plants appear and early terrestrial ecosystems are established.
Major TaxaPlants“Throughout Silurian times, only small rhyniophytes (or rhyniophytoids) and lycophytes are known . Remarkably, even in these early phases of their evolution, plants had an almost worldwide distribution with records from North and South America, Europe, Africa, central Asia, China and Australia, presumably reflecting the wide dispersal potential of their spores and the lack of any competition. The global composition of these floras also appears to be fairly uniform, although this may in part be due to the problems of differentiating biological species within the plexus of these morphologically very simple plants” (Cleal & Thomas 2009, p. 203). The first major diversification of land plant life, such as rhyniophytes, zosterophylls, drepanophycaleans, lycophytes, trimerophytes, and (in the southern hemisphere) Baragwanathia (Fig. 2, right), began as early as Late Ludlow (Late Silurian) in Australia through Gedinnian (Early Devonian) on Laurentia. Plants of this “rhyniophytoid phase” were small vascular plants having an axial organisation and terminal sporangia such as Cooksonia. “Cooksonia, the early or even primordial land plant, apparently played a key role: it has been described from a number of regions, ranges widely from the Silurian to the early Devonian, and is represented by several species such as C. pertoni, C. paranensis, C. banksii, and doubtfully C. cambrensis, C. hemisphaerica (Gonez and Gerrienne 2010a) with many other specimens having been described in open nomenclature. Cooksonia also occupies a special position in the colonization of terrestrial habitats, which is a major aspect of early plant research based on broad studies of associations, their successions and distributions (e.g. Edwards & Wellman 2001; Kenrick et al. 2012). This complex paleoecological approach has attracted considerable attention over the past two decades” Kraft et al. 2019, p. 144). Kraft et al. 2019 envisage an “Initial Plant Diversification and Dispersal Event” to be one of the key steps in global terrestrialization. This first significant diversification and dispersal episode of vascular plants, which occurred in the PrÚdolÚ (latest Silurian) in several paleoregions representing wide paleolatitude and paleoclimatic ranges, such as Avalonia, Laurentia, Baltica, Gondwana, South and North China and Kazakhstania (e.g., Edwards & Wellman 2001, Wellman et al. 2013, and other references in Kraft et al. 2019). |
(2)  and Baragwanathia; after Kraft et al. 2019, fig. 4.") Fig. 2: Cooksonia cf. hemisphaerica, left, and Baragwanathia brevifolia (Kraft et al. 2019, fig. 4). |
||||||||||
|
psilophytes – small Mid. Sil plants with horizontal rhizomes giving rise to vertical stalks bearing spore sacs ascomycete fungi Foraminifera were mostly agglutinated forms. Receptaculitids are an extinct group of organisms whose affinities are uncertain, though the majority view seems to favour placement within the algae. “Some paleontologists have interpreted them as a type of algae (that is, a sea-dwelling plant), and other paleontologists have considered them to represent a sponge-like animal. The receptaculitid skeleton is made of spiral rows of small attached plates. Receptaculitids were a minor group of dwellers in the Silurian reefs of Wisconsin” (taken from an old page, apparently removed now, on the Milwaukee Public Museum web site). Animals‘bounce-back’ following end-Ordovician extinctions Stromatoporoids – reef-building sponges first occurrence of coral reefs: large tabulate reefs common eurypterids common; reach their maximum diversity in Sil; some almost 2 metres long (very large for an exoskeleton) The earliest terrestrial body fossils are known from Silurian sediments: unassignable fragments from the Llandovery (Lower Silurian) age Tuscarora Formation in Pennslyvania, and identifiable arthropod fragments from the Late Silurian. widespread radiation of crinoids marine fishes diversify; first occurence in freshwater environments; wide and rapid spread of jawless fishes – ostracoderms – jawless armoured fish; jaws probably evolved from gill arch supports; first jawed fishes were acanthodians – jaws, paired fins, scales replace bony plates, spiney The Porifera are most notably represented by the Stromatoporoids, an extinct group of sponges that formed a compact, layered, calcareous skeleton. Stromatoporoids are known from Cambrian to Oligocene marine sediments, becoming dominant in the Silurian and Devonian. Like modern sponges, they appear to have been filter-feeders, actively pumping water to extract food particles. The stromatoporoid skeleton somewhat resembles compound tabulate corals in mostly adopting a generally dome-like form, or else thin flat sheets, often quite large. Stromatoporoids were a major type of reef constructor. (Read more: Page about stromatoporoids from University of California at Berkeley) Tetracorals (rugose corals) and tabulate corals were very common; the Silurian saw the first occurrence of coral reefs. Larger tabulate corals are found in coral-stromatoporoid reefs and, though important, they were not the frame-builders since they lacked any proper means of attachment. Smaller Ordovician and Silurian tabulates frequently occur in association with solitary rugose corals. Among the Mollusca, cephalopods were important – specifically the nautiloids (subclass Nautiloidea), which are characterised by simple suture lines, a non-calcareous protoconch, and umbilical perforation under the shell mantle, were most abundant in the Ordovician and Silurian. Only a few possessed planispiral shells, like that of their sole living representative, Nautilus: the majority were of either orthocone or cyrtocone (straight or slightly curved cones) shape. It is often assumed that the shells were entirely enclosed the body but there is good evidence that at least some forms possessed living tissue, including lateral fins, external to the shell. It is possibly from some of these taxa that the later belemnites were derived. |
|||||||||||
|
Trilobites were greatly reduced in numbers and on the whole display little more specialisation than earlier forms. The Ordovician trilobites were more successful at exploiting new environments, notably reefs, but during the mass extinction at the end of the Ordovician, they too suffered a crisis. Some distinctive and previously successful forms such as the Trinucleoidea and Agnostoidea became extinct.
Whiterock Fauna families, and in particular Silurian Fauna groups, can generally be traced only to the Early Ordovician, and in many cases they are entirely “cryptogenetic.” These clades certainly had Cambrian forebears, but the fact that they have avoided detection is a strong indication that novel morphologies were being developed very rapidly. Of the Silurian Fauna genera present during the Llandovery epoch, 78% also originated in that epoch, which demonstrates that extinctions were followed by a rapid post-extinction “rebound.” However, once this Llandovery rebound was completed, standing diversity returned to and was maintained at pre-extinction amounts. (After Adrain et al. 1998.) ‘Dudley Bug’ = Calymene blumenbachii; incorporated into the town’s coat of arms Silurian and Devonian assemblages are superficially similar, dominated by Lichiida and Phacopida, the latter including the well-known Calymenina. However, a number of characteristic forms do not extend far into the Devonian and almost all the remainder were wiped out by a series of extinctions in the Middle and Late Devonian. Only the Proetida survived the Devonian, lasting through the Carboniferous and Permian periods up until the Permo-Triassic mass extinction event. |
INSERT FIGURES – 2X SILURIAN TRILOBITES | ||||||||||
|
Eurypterids, an extinct group of aquatic chelicerates, attained their maximum diversity in the Silurian. They “were undoubtedly affected by environmental constraints imposed by their physiology and gross morphology, as are modern aquatic organisms. Kjellesvig-Waering (1961) proposed a series of distinct ecological phases for eurypterids, defined by the environment in which they lived, and based on the Upper Silurian Welsh Borderland fauna. However, evidence from the eurypterid fauna of the Upper Silurian Bertie Waterlime Formation, New York, suggests that two distinct transitionary assemblages existed, perhaps caused by a difference in the environmental preferences of juvenile and adult eurypterids (‘ontogenetic segregation’) (Manning 1993). The eurypterids from the Upper Silurian of the Welsh Borderland may represent a previously undescribed, ontogenetically mixed, eurypterid assemblage, influenced by a series of facies changes; this is supported by sedimentological evidence (Manning 1993). The distinct ecological phase model (Kjellesvig-Waering 1961) is rejected in favour of ecological gradients overlapping non-distinct eurypterid phases (inter-phase mixing). Inter-phase mixing might have been complicated further by the influence of ontogenetic migration of species across both physiological and environmental gradients. Combining this with new fossil evidence on the dual respiratory and osmoregulatory systems of eurypterids (Manning 1993; Manning & Dunlop 1995) leads to a better understanding of the palaeoecology of this enigmatic group” (Manning 1999). terrestrial arthropods: The earliest terrestrial body fossils are unassignable fragments from the Llandovery (Lower Silurian) Tuscarora Formation near Millerstown, Pennslyvania. Acid digestion of rock samples has recovered pieces of bristles or setae which may be arthropodan although they do not appear to be hollow, as are nearly all arthropod setae. Annelid setae are not hollow (Gray & Boucot 1994). The earliest identifiable terrestrial body fossils are arthropod fragments from the Late Silurian. However, these organisms possessed sensory and respiratory structures fully adapted for life on land, indicating an earlier history of terrestrial habituation that has not yet been found. ‘Bryozoans’ (Ectoprocta) were extremely abundant and diversified. “[T]he Wenlock bryozoans remain poorly studied and understood, even though they constitute an important part of the fauna. Species concepts are often insufficient, and there is a need for revised descriptions, incorporating thin-section work. Furthermore, phylogenetic, ecological and functional morphological studies also need to be carried out, as these areas have not been investigated in detail. Preliminary work on one particularly characteristic Wenlock bryozoan, Ptilodictya lanceolata (Goldfuss 1829), a cryptostome, has revealed new information on its functional morphology, together with patterns of seasonal development based on zooid measurements. In addition, multivariate statistical analyses are currently being used to review the fenestrate species. Past workers have relied too heavily on the so-called ‘micrometric formula’ and, as a result, taxonomically important characters have often been overlooked, and little thin-section work has been carried out. It is hoped that this research, utilising a wider range of characters, will provide a reliable taxonomic platform for the study of Silurian bryozoan phylogeny and palaeoecology” (Snell 1999). Brachiopods continued to diversify in the Silurian; the first spirifers appear continued diversification of brachiopods Echinodermata “The Much Wenlock Limestone Formation [has yielded] well over 600 species of invertebrates recorded. The Crinoidea account for around 10% of this number with an estimated 35 genera and 56 species. The high quality of preservation has revealed a range of small-scale morphological features among the aboral surfaces of the arms and calyces of this group. Small, circular to sub-circular depressions and parabolic traces are randomly situated in the calcite plates of a number of the crinoid taxa. A small number of these features indicate in vivo formation through the presence of rims and gall-like features surrounding the trace. These structures are interpreted as a response by the crinoid to the presence of another organism through either a mechanical or a chemical stimulant. The lack of penetration into the body cavity and the extent of the reaction structures suggest a symbiotic relationship existed between host crinoid and trace maker. A high degree of host-selectivity is observed with only 15% of crinoid species being affected. This number includes some of the most abundant crinoid species in fossil collections as well as a number of much rarer species. Work is now commencing on the taxonomic identity of the pit-producing organisms” (Widdison 1999). Graptolithina Craniata - conodonts common Marine fishes diversified rapidly in the Silurian, and here also we find their first occurences in freshwater environments. There was a wide and rapid spread of jawless fishes – ostracoderms – jawless armoured fish. The heterostracans – a diverse group of ancient, jawless fishes – underwent their major radiation in the Silurian, becoming common in Laurussia throughout the Devonian. Another jawless group, the Anapsida, forerunners of today’s lampreys, also flourished during the Silurian and early part of the Devonian. Jaws probably evolved from the gill arch supports of jawless fishes in the Ordovician, though it is in the Silurian that we begin to find widespread fossils of craniates with jaws. Acanthodians, an unusual group of mostly small, jawed fishes which first appear in the Early Silurian sediments of China, are unique among fishes in having ornamented bony spines in front of all their fins, and tiny scales with swollen, bulbous bases. Their affinities are unclear but seem to lie with the higher jawed fishes such as placoderms and osteichthyans. The bizarre, armour-plated placoderms also first appear in the Silurian, about 420 Ma. One group, the dinichthyids, achieved lengths of 8 m or more, the first creatures ever to reach such large sizes. A famous member of this group is Dunkleosteus. Finally, the bony fishes, which today form the largest and most diverse group of all vertebrates, the osteichthyans, arose about 410 Ma, in the latest Silurian or possibly earliest Devonian. Within a few million years all major groups of bony fishes had emerged and, before the end of the Devonian, some of them had crawled out of the water and begun to invade the land. (After Long 1995.) LagerstðttenLagerstätten (sing. Lagerstätte) are fossil localities which are highly remarkable for for either their diversity or quality of preservation; sometimes both. Some of the several famous lagerstätten of Silurian age are briefly described below.Wenlock Limestone: The Much Wenlock Limestone Formation of Wales and the Welsh Borderland preserves one of the most diverse, and well-preserved fossil assemblages known. More than 600 species of invertebrates have been recorded, including crinoids, corals, brachiopods, trilobites, algae and bryozoans. The formation was deposited during the early Silurian when this area of Britain was covered by a relatively warm, shallow shelf sea. Fiddler’s Green Formation: – 425 Ma; a formation within the Bertie Group; exposures in eastern New York State are particularly famous for their excellent eurypterid faunas Waukesha: – Wisconsin; arthropods and conodont animals; Mikulic et al. 1985. Lesmahagow: – Scotland; arthropods and fish; Ritchie 1985
New Zealand OccurrencesFor most of the Paleozoic and Mesozoic, the rocks which would become the basement rocks of the Zealandia continent formed part of the Pacific margin of Gondwana, flanking Australia and Antarctica (Edbrooke 2017; Strogen et al. 2017). During this time, the Zealandia basement developed mainly by subduction-driven, episodic accretion. The Silurian is poorly represented in New Zealand. Silurian fossils, mostly brachiopods, have been reported from only two or three localities in northwest Nelson, all in the Takaka Tarrane (Cooper 2004, p. 55). ReferencesAdrain, J.M.; Fortey, R.A.; Westrop, S.R. 1998: Post-Cambrian trilobite diversity and evolutionary faunas. Science 280: 1922-1925. Chaloner, W.G.; Lawson J.D. (ed.) 1985: Evolution and Environment in the Late Silurian and Early Devonian. Philisophical Transactions of the Royal Society of London, Series B 309: 1-342. Cleal, C.J.; Thomas, B.A. 2009: An Introduction to Plant Fossils. Cambridge University Press: 1-248. Cohen, K.M.; Finney, S.C.; Gibbard, P.L.; Fan, J.X. 2015: The ICS international chronostratigraphic chart v 2015/01. Episodes 36: 199-204. Cooper, R.A. (ed.) 2004: The New Zealand geologic timescale. Institute of Geological and Nuclear Sciences Monograph 22: 1-284. Edbrooke, S.W. 2017: The geological map of New Zealand. GNS Science Geological Map 2: 1-183. Edwards, D.; Wellman, C.H. 2001: Embryophytes on land: the Ordovician to Lochkovian (Lower Devonian) record. In Gensel, P.G.; Edwards, D. (ed.) 2001: Plants Invade the Land. Evolutionary and Environmental Perspectives. Columbia University Press: 3-28. Goldfuss, A. 1829: Petrefacta Germaniae. Düsseldorf 1: 1-128. Gonez, P.; Gerrienne, P. 2010a: A new definition and a lectotypification of the genus Cooksonia Lang 1937. Int. J. Plant Sci. 171 (2): 199-215. Gray & Boucot 1994: Early Silurian Nonmarine Animal Remains and the Nature of the Early Continental Ecosystem. Acta Palaeontol. Pol. 38: 303-328. Hamilton, B. 2001: Charles Lapworth's "The Moffat Series", 1878. Episodes 24 (3): 194-200. Kenrick, P.; Wellman, C.H.; Schneider, H.; Edgecombe, G.D. 2012: A timeline for terrestrialization: consequences for the carbon cycle in the Palaeozoic. Philisophical Transactions of the Royal Society of London, Series B 367 (1588): 519-536. Kjellesvig-Waering, E. N. 1961: The Silurian Eurypterida of the Welsh Borderland. Journal of Paleontology 35: 784-835. Kraft, P.; Psenicka, J.; Sakala, J.; Frýda, J. 2019: Initial plant diversification and dispersal event in upper Silurian of the Prague Basin. Palaeogeography, Palaeoclimatology, Palaeoecology 514 (2019) 144-155. Lapworth, C. 1878: The Moffat Series. Quarterly Journal of the Geological Society of London 34: 240-346. — 1879: The Silurian age. MS Lecture notes: University of Birmingham Archives. Long, J.A. 1995: The Rise of Fishes. Johns Hopkins. Manning, P.L. 1993: Palaeoecology of the eurypterids of the Upper Silurian of the Welsh Borderland. Unpublished M.Sc. thesis, University of Manchester. — 1999: Palaeoecology of Eurypterids from the Upper Silurian of the Welsh Borderland. Oral presentation to 43nd Annual Meeting, Palaeontological Association, University of Manchester, 19-22 December 1999. Manning, P.L.; Dunlop, J. 1995: The Respiratory Organs of Eurypterids. Palaeontology 38: 287-297. Mikulic, D.G.; Briggs, D.E.G.; J. Kluessendorf, J. 1985: A Silurian soft-bodied biota. Science 228: 715-717. Murchison, R.I. 1835: On the Silurian System of rocks. London and Edinburgh Philosophical Magazine and Journal of Science 7: 46-52. Ogg, J.G.; Ogg, G.; Gradstein, F.M. 2008: The concise geologic time scale. Cambridge University Press: 1-177. Ritchie, A. 1985: Ainiktozoon loganese Scourfield, a protochordate from the Silurian of Scotland. Alcheringa 9: 115-142. Snell, J. 1999: Wenlock Bryozoa from Wales and the Welsh Borderland. Poster presentation to 43nd Annual Meeting, Palaeontological Association, University of Manchester, 19-22 December 1999. Strogen, D.P.; Seebeck, H.; Nicol, A.; King, P.R. 2017: Two-phase CretaceousPaleocene rifting in the Taranaki Basin region, New Zealand; implications for Gondwana breakup. Journal of the Geological Society, London 174: 929-946. Wellman, C.H.; Steemans, P.; Vecoli, M. 2013: Palaeophytogeography of Ordovician-Silurian land plants. In Harper, D.; Servais, T. (ed.) 2013: Early Palaeozoic Biogeography and Palaeogeography. Geological Society London Memoirs 38: 461-476. Widdison, R. 1999: Biotic Interactions in Wenlock Crinoidea. Oral presentation to 43nd Annual Meeting, Palaeontological Association, University of Manchester, 19-22 December 1999. |
| Peripatus Home Page |
Hits counted from 27 Jan 2017:
My Traffic Estimate