| Peripatus Home Page | Updated: 07-Feb-2024 |
AbstractThis page describes the Carboniferous Period, including stratigraphy, paleogeography, and famous lagerstðtten, followed by a sketched outline of some of the major evolutionary events.Keywords: Carboniferous, Carboniferous biota, fossil record, evolution IntroductionThe Carboniferous was the first system to receive a name that is still in official use today. The name was applied to coal-bearing rocks in England and Wales by William Conybeare and William Phillips in 1822. Subdivisions of the Carboniferous have varied over time and between regions: In eastern Europe and Asia, Lower, Middle and Upper divisions were commonly used; in western Europe, just Lower and Upper; and, in North America, “Carboniferous” was typically not used at all, being replaced with Mississippian (lower) and Pennsylvanian (upper) Systems instead. The current international convention is to recognise the Carboniferous System (Period) as being divided into Mississippian and Pennsylvanian Series (Epochs). (After Ogg et al. 2008, p. 73.) |
|
||||||||||
StratigraphyHistorical Development
Lower (Devonian–Carboniferous) BoundaryThe GSSP defining the the base of the Carboniferous (i.e., the Devonian-Carboniferous Boundary) was established in 1991 (Paproth et al. 1991) at La Serre, France, where it is set at the first appearance datum (FAD) of the conodont Siphonodella sulcata. Subsequent research, however, has revealed stratigraphic problems with this datum, and it is now relised that this horizon cannot be correlated with precision at present. The GSSP is at the base of Bed 89 in Trench E at La Serre. The La Serre section is located in the southeastern Montagne Noire, Departement HÕrault, District of CabriÒres, in southern France. Artificial trench E, which averages 80cm in depth on the southern slope of La Serre Hill, is about 125m south of the hilltop (252m), about 525m east of La Roquette farmhouse, 2.5km northeast of the village of FontÒs (sheet 1:25,000 PÕzenas XXVI/44, 1-2, x = 682.55, y = 140.12). Sedimentology: The rocks in trench E’ are part of a geologic unit named “Klippen of CabriÒres”. The youngest Devonian and earliest Carboniferous beds are characterized by a sequence of predominantly biodetrital oolitic limestone within a pelagic matrix of shale and cephalopod bearing calcilutites. Primary Markers: First appearance of the conodont Siphonodella sulcata within the evolutionary lineage from Siphonodella praesulcata to Siphonodella sulcata at the base of Bed 89 in trench E’. Secondary Markers: The trilobites Belgibole abruptirhachis, Archegonus (Phillibole) and Carbonocoryphe also occur in Bed 89. Some of these other taxa are especially important at other Locations around the world. For example, Belgibole abruptirhachis occurs immediately above the Hangenberg Schiefer interval in various cephalopod-bearing sections from the Renish Slate Mountains (Germany), the Holy Cross Mountains (Poland) and the Carnic Alps (Austria). See Paproth et al. 1991.
Upper (Carboniferous–Permian) BoundaryThe GSSP defining the the base of the Permian (i.e., the Carboniferous-Permian Boundary) was established in 1998 (Davydov et al. 1998) at Aidaralash Creek, Kazakhstan, at the first appearance of the “isolated-nodular” morphotype of the conodont Streptognathodus “wabaunsensis”.
ChronologyCurrent age calibrations place the base of the Carboniferous and Mississippian at 358.9 ±0.4 Ma, the base of the Pennsylvanian at 323.2 ±0.4 Ma, and the base of the Permian (i.e., the top of the Carboniferous and Pennsylvanian) at 298.9 ±0.15 Ma (Cohen et al. 2015). Thus the Mississippian is approximately 36 my in duration, the Pennsylvanian approximately 24 my, and the whole Carboniferous Period approximately 60 my. PaleogeographyMajor Tectonic EventsMajor episodes of mountain building were associated with the assembly of Pangea. Both the Appalachians and the precursor ranges of the Rocky Mountains in North America began forming at this time. |
|||||||||||
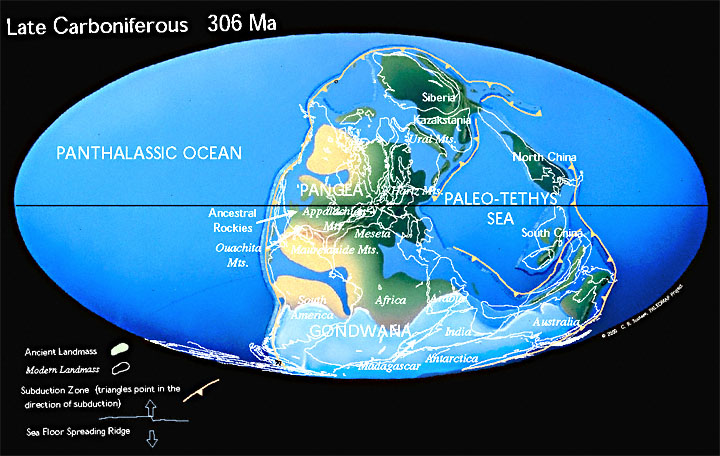
Land and SeaAt the beginning of the Carboniferous Period, the supercontinent of Pangea was forming from the collision of Laurentia (North america and Europe) and northern Gondwana (Fig. 1). “Before the collision, Gondwana had rotated clockwise, so that the its eastern part (India, Australia, and Antarctica) moved south while its western part (South America and Africa) moved north. This rotation opened a new ocean, the Tethys Ocean, in the east and closed an old one, the Rheic, in the west” (Palmer 1999, p. 26).The assembly of Pangea concentrated almost the entire landmass of the Earth into one hemisphere; the Panthallassic Ocean occupied almost the entire opposite hemisphere. In general, sea levels rose through the early part of the Carboniferous, receding again in the later part. |
(1) |
||||||||||
ClimateThe global climate cooled from the humid, hothouse conditions of the preceding Devonian Period. By the mid-Carboniferous it had become markedly seasonal and polar icecaps were forming. By the end of the period, aridity had increased and glaciation had become extensive.Atmospheric oxygen increased to perhaps 35% towards the end of the Period. Carbon dioxide levels were also high by today’s standards. PaleontologyIntroductionThe seas were dominated by sharks and bony (teleost) fishes. On land, arthropods such as Arthropleura and the early griffenflies became very large, and tetrapods diversified.
Major TaxaPlants“In Pennsylvanian [= Late Carboniferous] times, there was a dramatic change in vegetation. In low paleolatitudes, the very first tropical rain forests appeared: the so-called ‘Coal Forests’ of europe, North America and China (known as the Amerosinian Palaeokingdom...), so named because of the vast reserves of coal-forming peat that were laid down by them . These forests were dominated by the giant lycophytes , which were perfectly adapted to the wetland habitats that existed over much of the then tropics” (Cleal & Thomas 2009, p. 205-206). Tetrapods“The development of limbs from fins was a defining stage in the evolution of tetrapods (Clack 2002a). The earliest limbs with digits are thought to have been used to facilitate underwater bottom walking (Coates & Clack 1995), but the critical next step was their use in terrestrial locomotion. [However,] the evolution of limbs enabling tetrapods to use quadrupedal gaits on land is still poorly understood” (Smithson & Clack 2018, p. 89). “The fossil record of early tetrapods has been increased recently by new finds from the Devonian period and mid–late Early Carboniferous period. Despite this, understanding of tetrapod evolution has been hampered by a 20-million-year gap (‘Romer’s Gap’) that covers the crucial, early period when many key features of terrestrial tetrapods were acquired” (Clack 2002b, p. 72). “This period between the end Devonian and the mid-Viséan represents the time when tetrapods underwent a major diversification and acquired true terrestriality” (Clack & Finney 1999). Romer’s Gap is an apparent gap in the tetrapod fossil record (or, more generally, the record of any terrestrial animal; see Ward et al. 2006) extending through the early part of the Carboniferous, lasting for up to 20 million years following the end-Devonian extinction. The name appears to have first been coined by Coates & Clack (1995). “The first half of the Mississippian or Early Carboniferous (Tournaisian to mid-Viséan), an interval of about 20 million years, has become known as ‘Romer’s Gap’ because of its poor tetrapod record. Recent discoveries emphasise the differences between pre-’Gap’ Devonian tetrapods, unambiguous stem-group members retaining numerous ‘fish’ characteristics indicative of an at least partially aquatic lifestyle, and post-’Gap’ Carboniferous tetrapods, which are far more diverse and include fully terrestrial representatives of the main crown-group lineages. It seems that ‘Romer’s Gap’ coincided with the cladogenetic events leading to the origin of the tetrapod crown group” (Chen et al. 2018, Abstract). Sallan & Coates (2010) explain Romer’s Gap as the post-extinction trough following the Hangenberg extinction at the end of the Devonian. “Corresponding lack of Tournaisian material from other terrestrial groups (e.g., insects) supports this conclusion..., and Devonian levels of faunal disparity are only regained in the Serpukhovian.... Even late Mississippian assemblages of vertebrate species from both sides of the newly formed Pangaea are strikingly similar (e.g., Bearsden and Bear Gulch localities...), likewise for associated invertebrate assemblages . Extinctions remove characters from the pool of varying morphologies.... For example, digit number is known to be variable among late Famennian [the last age in the Devonian Period] tetrapods ... but stabilizes with a maximum limit of five among all later forms” (Sallan & Coates 2010, p. 10134, 10135). The “gap”, however, has now been shown to be the result of collection failure (see Smithson et al. 2012, Clack et al. 2016, Smithson & Clack 2018). Notwithstanding, the only articulated skeleton of a tetrapod yet found from the Tournaisian epoch (354–344 Ma) is Pederpes, “the earliest-known tetrapod to show the beginnings of terrestrial locomotion and was at least functionally pentadactyl” (Clack 2002b, Abstract). With its later American sister-genus, Whatcheeria, it represents the next most primitive tetrapod clade after those of the Late Devonian, bridging the temporal, morphological and phylogenetic gaps that have hitherto separated Late Devonian and mid-Carboniferous tetrapod faunas” (Clack 2002b, p. 72). Chen et al. (2018) describe a partial right jaw ramus of a new tetrapod, Tantallognathus woodi, from the late Tournaisian or early Viséan of Scotland. The large and robust jaw exhibits a distinctive combination of characters. A phylogenetic analysis places Tantallognathus in the upper part of the tetrapod stem group, above Pederpes and Whatcheeria. The earliest terrestrial vertebrates are often informally referred to as “amphibians”. “Traditionally the name ‘Amphibia’ has been employed more broadly to refer to all tetrapods that are not amniotes, but this is rather misleading because most of these forms were not similar to extant amphibians and may have been different biologically. Among the Paleozoic and early Mesozoic ‘amphibians,’ most authors have distinguished two principal groups – Lepospondyli and Temnospondylli. The exclusively Paleozoic lepospondyls comprise a heterogeneous assemblage of small forms with holospondylous vertebrae. By contrast, temnospondyls are a remarkably diverse group that ranged from the Mississippian to the Early Cretaceous and include the stem forms of at least frogs and salamanders. During the Triassic, temnospondyls were represented by a variety of lineages and were semi-aquatic or fully aquatic carnivores in many ecosystems. Most Triassic temnospondyls are referred to the Stereospondlyi,” a subgroup of which is the Capitosauroidea. Capitosauroids “include often large forms with a dorsoventrally flattened truck and head, and an elongated snout, which lends them a superficially crocodile-like appearance. A characteristic feature is the presence of a deeply incised otic notch, or opening (which presumably accommodated the tympanic membrane), along the posterior margin of the skull roof” (Sues & Fraser 2010, p. 12). Beyond evolving limbs capable of terrestrial locomotion, later tetrapods evolved the amniote egg, thereby becoming truly terrestrial, although it is not known exactly when. Most reptiles have soft parchment or membrane-like eggshells which are much less likely to fossilise than calcareous eggshells (Hirsch 1979, p. 1068). The oldest known fossil which might plausibly be an egg is Early Permian. The presumtive fossil egg was found by Llewellyn Price, having weathered free from the upper part of the Admiral Formation, Witchita Group, in the wonderfully-named Rattlesnake Canyon, Archer County, Texas. Romer & Price were unsure of the age at that time, whether latest Carboniferous or Early Permian, but the age was later established as Early Permian (Romer 1974). The fossil itself is small – about 59 mm long by 36 diameter at its widest point – asymmetrically ovoid, somewhat blunt at both ends. The surface is studded irregularly with small, less than 1 mm, rounded swellings. It was first described by Romer & Price (1939) and re-examined some 40 years later by Hirsch (1979) who removed small pieces for sectioning and compositional analysis. Hirsch noted that the outer “shell-like layer is not as uniform as described by Romer & Price and, contrary to their findings, thin wavy layers are visible throughout the specimen, at least to the depth to which the deepest cut was made (4.5 mm) . Although the colored wavy layers which are visible to the naked eye can be seen in the polarized photo taken under the metallograph ... they are not discernible on the SEM. Thus it seems that these layers are not structurally significant but are rather colored or stained bands of the matrix. ... Closer observation of the shell-like layer shows very little similarity to calcified fossil eggshell” (p. 1071, 1075). But although he emphatically rejected the idea of a calcareous eggshell, Hirsch seems to have remained open to the idea that the fossil might still be a soft-shelled egg: “Phosphorous is relatively high and constant in the outer layers and seems to be less common toward the inside of the specimen. This phosphorous might be the residue of a soft, parchment-like organic shell layer. ¶ Unfortunately, the described specimen could only be compared with one nodular specimen of the same formation. This comparison showed that node-like structures are also found on nodules and confirmed the assumption that the phosphorous in the Romer and Price specimen is not a common occurrence” (Hirsch 1979, p. 1083). To the best of my knowledge there has not been any further study. Four common animals are known from around the same stratigraphic horizon in the area: Diadectes, Dimetrodon, Edaphosaurus, and Ophiacodon. Romer & Price (1939, p. 829) note that the last of these is particularly abundant and “somewhat” favour it as the possible egg-layer. LagerstðttenMazon Creek: Late Carboniferous Mazon Creek Illinois; deltaic and near-shore marine; 318 MaBear Gulch: Late Early Carboniferous Bear Gulch Montana; estuarine; notable for fish and arthropod fossils Joggins: Nova Scotia; includes representatives of one of the oldest known reptils; Grey & Finkel 2011 Loch Humphrey Burn: Early Carboniferous Loch Humphrey Burn Southern Scotland; an exceptionally well-preserved Lower Carboniferous terrestrial ecosystem, containing several successive plant bearing horizons within a volcanic terrain. ‘Shrimp Beds’: Early Carboniferous Scottish ‘Shrimp Beds’ Scotland; crustaceans, conodont animals, tomopterid worms, fish; Briggs & Clarkson 1983, 1985, Briggs et al. 1991 East Kirkton: Early Carboniferous East Kirkton Scotland; hot spring deposits; plants; arthropods (scorpions); amphibians and reptiles; Rolfe 1988, 1994
New Zealand OccurrencesFor most of the Paleozoic and Mesozoic, the rocks which would become the basement rocks of the Zealandia continent formed part of the Pacific margin of Gondwana, flanking Australia and Antarctica (Edbrooke 2017; Strogen et al. 2017). During this time, the Zealandia basement developed mainly by subduction-driven, episodic accretion. “In an initial period of growth, from Cambrian to Carboniferous time, the oldest sedimentary rocks known in mainland New Zealand were deposited and acreted . Intrusion of granitic rocks occurred intermittently but mainly during Late Devonian and Early Carboniferous time” (Edbrooke 2017, p. 31). Carboniferous rocks are almost unknown in New Zealand. Only four isolated conodont faunas having been reported (Campbell in Cooper 2004, p. 61), accompanied in one case by rare fish scales (Jenkins & Jenkins 1971). ReferencesBriggs, D.E.G.; Clark, N.D.L.; Clarkson, E.N.K. 1991: The Granton shrimp-bed, Edinburgh – a Lower Carboniferous Konservat-Lagerstätte. Transactions of the Royal Society of Edinburgh, Earth Sciences 82: 65-86. Briggs, D.E.G.; Clarkson, E.N.K. 1983: The Lower Carboniferous Granton Shrimp-Bed Edinburgh. Special Papers in Palaeontology 30: 161-177. — 1985: The Lower Carboniferous Shrimp Tealliocaris from Gullane, East Lothian, Scotland. Transactions of the Royal Society of Edinburgh, Earth Sciences 76: 173-201. Chen, D.; Alavi, Y.; Brazeau, M.D.; Blom, H.; Millward, D.; Ahlberg, P.E. 2018: A partial lower jaw of a tetrapod from “Romer’s Gap”. Earth and Environmental Sciences Transactions of the Royal Society of Edinburgh 108: 55-65. Clack, J.A. 2002: Gaining ground: The Origin and Early Evolution of Tetrapods. Indiana University Press: 1-400. — 2002: An early tetrapod from ‘Romer’s Gap’. Nature 418: 72-76. Clack, J.A.; Bennett, C.E.; Carpenter, D.K.; Davies, S.J.; Fraser, N.C.; Kearsey, T.I.; Marshall, J.E.A.; Millward, D.; Otoo, B.K.A.; Reeves, E.J.; Ross, A.J.; Ruta, M.; Smithson, K.Z.; Smithson, T.R.; Walsh, S.A. 2016: Phylogenetic and environmental diversity revealed for Tournaisian tetrapods. Nature Ecology and Evolution 1: 1-11. Clack, J.A.; Finney, S.M. 1999: The First Articulated Tournaisian Tetrapod: an Update. Palaeontological Association 43rd Annual Meeting, University of Manchester, 19-22 December 1999 (Poster Presentation). Cleal, C.J.; Thomas, B.A. 2009: An Introduction to Plant Fossils. Cambridge University Press: 1-248. Coates, M.I.; Clack, J.A. 1995: Romer’s Gap - tetrapod origins and terrestriality. Bulletin du Muséum national d’Histoire naturelle 17: 373-388. Cohen, K.M.; Finney, S.C.; Gibbard, P.L.; Fan, J.X. 2015: The ICS international chronostratigraphic chart v 2015/01. Episodes 36: 199-204. Conybeare,W.D.; Phillips,W. 1822: Outlines of the geology of England and Wales, with an introduction compendium of the general principles of that science, and comparative views of the structure of foreign countries. Part 1. William Phillips, London: 1-563. Cooper, R.A. (ed.) 2004: The New Zealand geologic timescale. Institute of Geological and Nuclear Sciences Monograph 22: 1-284. Davydov, V.I.; Glenister, B.F.; Spinosa, C.; Ritter, S.M.; Chernykh, V.V.; Wardlaw, B.R.; Snyder, W.S. 1998: Proposal of Aidaralash as Global Stratotype Section and Point (GSSP) for base of the Permian System. Episodes 21 (1): 11-18. Edbrooke, S.W. 2017: The geological map of New Zealand. GNS Science Geological Map 2: 1-183. Grey, M.; Finkel, Z.V. 2011: The Joggins Fossil Cliffs UNESCO World Heritage site: a review of recent research. Atlantic Geology 47: 185-200. Hirsch, K.F. 1979: The Oldest Vertebrate Egg? Journal of Paleontology 53 (5): 1068-1084. Jenkins, D.G.; Jenkins, B.H. 1971: First diagnostic Carboniferous fossils from New Zealand. Nature 233: 117-118. Ogg, J.G.; Ogg, G.; Gradstein, F.M. 2008: The concise geologic time scale. Cambridge University Press: 1-177. Palmer, D. 1999: Atlas of the prehistoric world. Marshall Publishing: 1-224. Paproth, E., Feist, R., and Flaijs, G. 1991: Decision on the Devonian-Carboniferous boundary stratotype. Episodes 14 (4): 331-336. Rolfe, W.D.I. 1988: Early life on land; the East Kirkton discoveries. Earth Science Conservation 25: 22-28. Rolfe, W.D.I.; Durant, G.P.; Baird, W.J.; et al. 1994: The East Kirkton Limestone, VisÕan, West Lothian, Scotland: an introduction and stratigraphy. Transactions of the Royal Society of Edinburgh, Earth Sciences 84: 177-188. Romer, A.S. 1974: The stratigraphy of the Permian Wichita redbeds of Texas. Brevioria 427: 1-31. Romer, A.S.; Price, L.I. 1939: The oldest vertebrate egg. American Journal of Science 237: 826-829. Sallan, L.C.; Coates, M.I. 2010: End-Devonian extinction and a bottleneck in the early evolution of modern jawed vertebrates. Proceedings of the National Academy of Sciences of the USA 107 (22): 10131-10135. Smithson, T.R.; Clack, J.A. 2018: A new tetrapod from Romer’s Gap reveals an early adaptation for walking. Transactions of the Royal Society of Edinburgh 108: 89-97. Smithson, T.R.; Wood, S.P.; Marshall, J.E.A.; Clack, J.A. 2012: Earliest Carboniferous tetrapod and arthropod faunas from Scotland populate Romer’s Gap. Proceedings of the National Academy of Science USA 109: 4532-4537. Strogen, D.P.; Seebeck, H.; Nicol, A.; King, P.R. 2017: Two-phase CretaceousPaleocene rifting in the Taranaki Basin region, New Zealand; implications for Gondwana breakup. Journal of the Geological Society, London 174: 929-946. Sues, H.-D.; Fraser, N.C. 2010: Triassic life on land. Columbia University Press: 1-236. Ward, P.; Labandeira, C.; Laurin, M.; Berner, R.A. 2006: Confirmation of Romer’s Gap as a low oxygen interval constraining the timing of initial arthropod and vertebrate terrestrialization. Proceedings of the National Academy of Sciences of the USA 103 (45): 16818-16822. |
| Peripatus Home Page |
Hits counted from 2 Jun 2017:
My Traffic Estimate